








Servicios Personalizados
Revista

Articulo
 Portugués (pdf)
Portugués (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versión impresa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.5 n.2 Belém ago. 2010
Tricomas fósseis como nova categoria de palinomorfos lato sensu: sua classificação preliminar e aplicabilidade na Bioestratigrafia
Fossil trichomes as a new category of palynomorphs sensu lato: their preliminary classification and applicability in biostratigraphy
Mitsuru AraiI; Sarah Gonçalves DuarteII
ICentro de Pesquisas da Petrobras. Rio de Janeiro, Rio de Janeiro, Brasil (arai@petrobras.com.br)
IIUniversidade Federal do Rio de Janeiro. Rio de Janeiro, Rio de Janeiro, Brasil (sarahpalino@yahoo.com.br)
RESUMO
Estruturas vegetais reconhecidas como tricomas (apêndices unicelulares ou pluricelulares encontrados na epiderme dos vegetais) vêm sendo registradas em estratos cretáceos submetidos a análises palinológicas. O objetivo deste trabalho é sistematizar o registro de tricomas fósseis, classificando-os sob um esquema taxonômico artificial aplicável na bioestratigrafia. O material analisado provém, em sua maior parte, de poços petrolíferos que perfuraram o Cretáceo das bacias de Campos e Santos (plataforma continental do Sudeste do Brasil). Com base em caracteres selecionados (número de ramificações, diâmetro relativo, número de células, tipo da base, ângulo entre as ramificações, forma do centro e pedicelo), foram identificados 12 morfotipos básicos: (1) simples; (2) em forma de T; (3) em forma de Y; (4) tipo duplo; (5) triplo; (6) fasciculado; (7) fasciculado tufoso; (8) multirradiado; (9) multirradiado tufoso; (10) estrelado tridimensional; (11) estrelado tridimensional tufoso; e (12) estrelado planar. Para cada um destes morfotipos, podem ser estabelecidos subtipos baseados em características secundárias (morfologia das extremidades das ramificações, curvatura, sinuosidade etc.), de modo que podem ser identificados 63 morfotáxons. Em função dessa grande variabilidade morfológica, tricomas acenam com potencial aplicabilidade na bioestratigrafia. Isto se torna especialmente importante, considerando que eles são frequentes dentro do Santoniano (Cretáceo Superior) das bacias de Santos e Campos, onde, em consequência de paleoincêndios provocados pelo vulcanismo, observa-se uma escassez de palinomorfos convencionais (esporos e grãos de pólen).
Palavras-chave: Brasil. Cretáceo. Palinologia. Tricomas. Bacia de Campos. Bacia de Santos.
ABSTRACT
Plant components recognized as trichomes (unicellular or multicellular structures occurring in the plant epidermis) have been being recorded in Cretaceous sediments submitted for palynological analyses. The objective of the present paper is to record the occurrence of these fossil trichomes, and to classify them by proposing an artificial scheme that could be applied for biostratigraphic purposes. The material analyzed derives mainly from petroleum wells that penetrated Cretaceous strata in the Santos and Campos basins of offshore southeastern Brazil. Based on selected characteristics (number of arms, relative diameter, number of cells, type of base, angle formed by arms, central area pattern and presence of stalk), twelve basic morphotypes are identifiable: (1) simple; (2) T-shaped; (3) Y-shaped; (4) double (twin); (5) triple; (6) fasciculate; (7) tufted fasciculate; (8) multiradiate; (9) tufted multiradiate; (10) three-dimensional stellate; (11) tufted three-dimensional stellate; and (12) planar (two-dimensional) stellate. Within each of these morphotypes, subtypes based on secondary characteristics (morphology of arm ends, curvature, sinuosity etc.) could be recognized, thus resulting in 63 morphotaxa. Clearly, this large morphological variability among trichomes presents significant scope for their application in biostratigraphy. This assumes especial importance in the Santonian (Upper Cretaceous) strata of the Santos and Campos basins, where paleowildfires caused by regional volcanism have resulted in the scarcity of conventional, biostratigraphically significant palynomorphs (spores and pollen grains).
Keywords: Brazil. Cretaceous. Palynology. Trichomes. Campos Basin. Santos Basin.
INTRODUÇÃO
Estruturas vegetais reconhecidas como tricomas - apêndices encontrados na epiderme das plantas (Fahn, 1990) - vêm sendo encontradas durante análises paleopalinológicas realizadas em apoio às atividades exploratórias da Petrobras. O objetivo do presente trabalho é tentar classificar artificialmente os morfotipos encontrados. O material analisado provém, em sua maior parte, de poços que perfuraram o Santoniano (85,8 - 83,5 Ma, Cretáceo Superior) das bacias de Campos e Santos (plataforma continental do Sudeste do Brasil) e, em menor parte, de afloramentos do Maastrichtiano (70,6 - 65,5 Ma, Cretáceo Superior) da Formação Urucutuca da bacia de Almada (bacia costeira da Bahia).
Nas lâminas palinológicas onde ocorre esse material, palinomorfos stricto sensu (esporos e grãos de pólen) são raros, em função do paleoincêndio regional que destruiu grande parte da matéria orgânica não enriquecida em lignina, fazendo com que predominem nelas esclereídes, tricomas e outros restos de matéria orgânica lenhosa carbonificada (Arai et al., 2006; Arai, 2008).
O presente trabalho segue a mesma linha da pesquisa já realizada com esclerócitos (esclereídes fósseis) por Duarte & Arai (2008, 2010), cuja finalidade prática é a busca de palinomorfos lato sensu utilizáveis ao zoneamento bioestratigráfico. O detalhamento bioestratigráfico do Santoniano da bacia de Santos reveste-se de caráter especial, pois o referido intervalo contém importantes reservatórios de petróleo e gás.
MATERIAL E MÉTODOS
Os dados de ocorrências de tricomas fósseis foram levantados a partir de 2008, concomitantemente às análises palinológicas de rotina do Centro de Pesquisas da Petrobras (CENPES). O material estudado é representado principalmente pelas amostras de calha e de testemunho provenientes dos poços exploratórios das bacias de Campos e Santos (Figura 1), em intervalos cretáceos de profundidade (cota abaixo do nível do mar) entre 3.300 m e 5.400 m. Foram analisadas também algumas amostras de afloramento do Maastrichtiano da bacia de Almada.
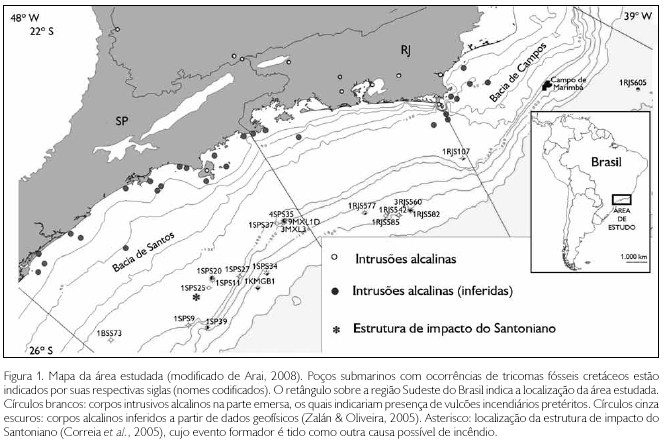
O método de preparação de amostras é o mesmo da Paleopalinologia tradicional, ou seja, processamento de amostras de rochas sedimentares com as macerações utilizando os ácidos clorídrico, fluorídrico e nítrico, seguido de montagem de lâminas palinológicas, que são analisadas ao microscópio óptico biológico (Uesugui, 1979). As fotomicrografias foram obtidas através de câmera digital Zeiss AxioCam MRc acoplada a microscópio Zeiss Axio Imager A1. Algumas amostras excepcionalmente ricas em tricomas foram analisadas também por microscópio eletrônico de varredura (MEV). Para isso, o resíduo palinológico, depois de seco, foi metalizado por ouro/paládio, em metalizador EMITECH modelo K575X, a fim de torná-lo condutivo. Em seguida, foram obtidas imagens por elétrons secundários ao MEV (ZEISS, modelo EVO 40) da Gerência de Bioestratigrafia e Paleoecologia do CENPES, operando com aceleração de voltagem de 20 kv.
HISTÓRIA NATURAL DOS TRICOMAS
De acordo com Fahn (1990), tricomas são apêndices encontrados na epiderme dos vegetais. Todavia, essa definição varia em diversos trabalhos de anatomia vegetal, podendo ter uma abordagem mais simples ou mais complexa, a depender da estrutura tecidual associada, origem tecidual, morfologia, função etc.
Os tricomas são encontrados desde pteridófitas até angiospermas, estando presentes em diversas partes das plantas, como nas superfícies de folhas, caules, raízes, flores e frutos. Podem ser unicelulares ou multicelulares, sendo que nestes, as células se organizam de diversas maneiras: alinhadas, ramificadas etc. (Fahn, 1990).
Alguns tricomas são glandulares, cujas funções são secreção de metabólitos em geral, aumento da superfície de absorção ou digestão e secreção de mucilagens para a captura de presas em plantas carnívoras, entre outras. Existe uma diversidade de substâncias secretadas pelos tricomas glandulares. Sais, mucos, essências, enzimas digestivas, gomas, soluções aquosas, néctares são apenas alguns de seus exemplos (Alquini et al., 2003). Os conteúdos de tricomas variam de acordo com suas funções, e os mais complexos são os de células glandulares (Netolitzky, 1932). Já os não glandulares ou tectores têm a função de tornar partes vegetais impalatáveis para alguns insetos, evitar a perda de água em algumas plantas xeromórficas ou controlar a perda de calor (Fahn, 1986; Fahn & Cutler, 1992).
Tricomas originam-se na epiderme de órgãos vegetais (Theobald et al., 1979). As paredes celulares deles são comumente constituídas de celulose e cobertas por uma cutícula, podendo ocasionalmente desenvolver paredes secundárias lignificadas. Em tricomas glandulares, podem ser encontradas estruturas de paredes com suberina e cutina. Alguns tricomas podem até perder seu protoplasto (Fahn, 1990).
Os tricomas, também chamados de pelos vegetais em alguns casos, produzem paredes secundárias espessas, como, por exemplo, no caso de pelos da semente de algodão (Anderson & Kerr, 1938). Existem também casos em que as paredes dos tricomas são impregnadas com sílica ou carbonato de cálcio (Beyrich, 1943). No interior de alguns tricomas simples, podem ocorrer cistólitos, que são estruturas "petrificadas" ricas em oxalato de cálcio e outros cristais (Esau, 1953).
Geralmente, tricomas impregnados de sílica e cálcio são os que servem de defesa para as plantas contra o ataque de parasitas. Essas plantas geralmente mobilizam estes elementos químicos dos sais, os quais são retirados por elas da água absorvida do solo, e deslocados para as paredes celulares dos tricomas ou das células epidérmicas (Esau, 1953).
Existem espécies de angiospermas que são completamente desprovidas de tricomas em todas as suas partes, todavia, elas representam uma minoria, geralmente decorrente de alguma mudança genética (Fahn, 1990). Tricomas têm sido utilizados como parâmetro de grande importância na investigação comparativa e sistemática de angiospermas, por estarem quase sempre presentes e serem facilmente observáveis, além de possuírem um variado padrão morfológico (Theobald et al., 1979).
Tricomas podem apresentar grande variação entre famílias, entre um pequeno grupo de plantas e, inclusive, dentro de uma mesma planta. Por outro lado, pode às vezes existir considerável uniformidade nos tricomas de um mesmo grupo vegetal. Eles têm sido usados com sucesso na classificação de gêneros e de espécies em certas famílias, assim com no reconhecimento de híbridos intraespecíficos (Metcalfe & Chalk, 1950).
Importantes estudos com tipos particulares de tricomas têm sido realizados em muitos grupos taxonômicos de angiospermas, como, por exemplo, nas famílias Gramineae (Metcalfe, 1960) e Asteraceae (Lodari et al., 1989; Sancho & Katinas, 2002; Freire et al., 2007), e em algumas ordens, como Commelinales e Zingiberales (Tomlinson, 1969).
Ainda existe muita dificuldade para o estudo e utilização de tricomas em análises taxonómicas comparativas, pelo fato dos termos utilizados atualmente em sua classificação não conseguirem expressar de maneira exata os verdadeiros aspectos morfológicos e anatômicos dos mesmos (Theobald et al., 1979).
CLASSIFICAÇÃO DOS TRICOMAS
Existem várias propostas de classificação dos tricomas. Payne (1978) classificou-os em doze categorias básicas, divididas entre tipo glandular e não glandular, considerando não só aspectos morfológicos, mas também aspectos funcionais. Já para Theobald et al. (1979), os tricomas podem ser classificados em sete categorias básicas (Figura 2), levando-se em conta somente as características morfológicas individuais dos mesmos: papiloso (papillae), simples (simple), ramificado (2-5 armed), estrelado (stellate), escamiforme (scales), dendrítico (dendritic) e tipos especializados. Estas categorias, por sua vez, podem se desdobrar em subcategorias.
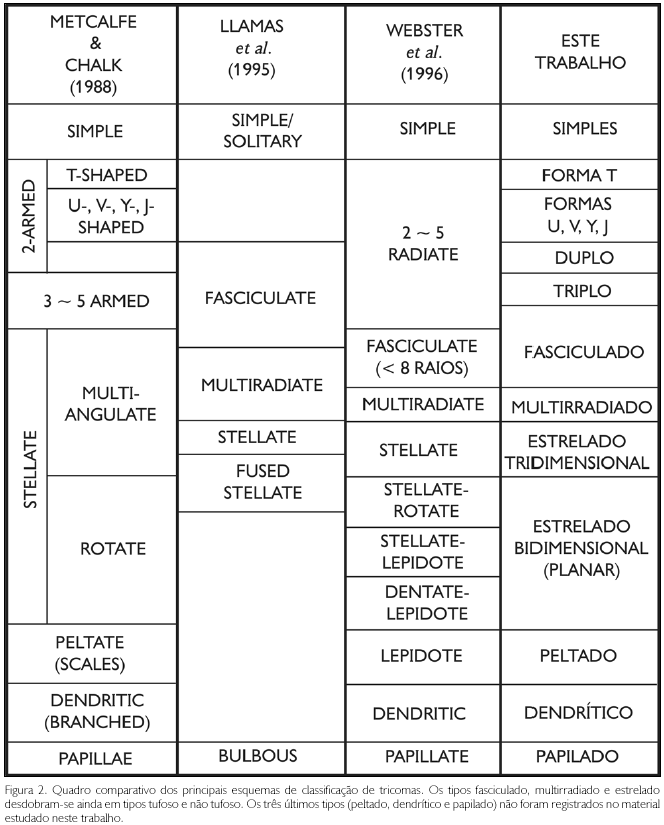
Outros autores classificaram tricomas dentro de um grupo taxonômico restrito. Llamas et al. (1995), ao estudar Quercus (Fagaceae), efetuaram a caracterização dos morfotipos fasciculado (fasciculate), multirradiado (multiradiate) e estrelado (stellate). Webster et al. (1996), ao estudar tricomas de Croton (Euphorbiaceae), detalharam a classificação dos morfotipos fasciculado (fasciculate), estrelado (stellate), peltado (lepidote) e dendrítico (dendritic).
O que fica evidente, ao analisar esses trabalhos, é o fato de que a mesma terminologia pode ser usada com sentidos diferentes. A comparação dos principais esquemas de classificação pode ser visualizada no Quadro Comparativo (Figura 2). Alguns autores admitiram que, embora houvesse progresso no estudo sobre tricomas nas últimas décadas, sua classificação definitiva está ainda longe de ser alcançada (Theobald et al., 1979). Para outros, a dificuldade na padronização da terminologia é justificada pela existência de imensa variabilidade morfológica (e.g., Webster et al., 1996).
PROCESSO DE FOSSILIZAçÃO DOS TRICOMAS
Tricomas são passíveis de serem fossilizados, pois, da mesma forma que as esclereídes, podem desenvolver paredes secundárias que, em alguns casos, tornam-se lignificadas.
A lignina é uma substância orgânica que resiste aos processos de preparação palinológica, principalmente se for submetida ao aquecimento, como aconteceu com a maioria dos materiais estudados no presente trabalho, os quais foram afetados pelo paleoincêndio antes de seu transporte e deposição (Arai et al., 2006; Arai, 2008). Entretanto, existem tricomas que provavelmente não se preservam nas lâminas palinológicas. Esta suspeita decorre da observação de Milanez & Machado (2008), que realizaram um estudo de anatomia foliar de Microlepis oleaefolia (DC.) Triana (Melastomataceae), no qual constataram que as extremidades dos tricomas são de composição péctica, ao passo que a base é mais lignínica, chegando mesmo a constituir esclereídes em suas raízes. Caso esses tricomas passassem pelo processo de fossilização, dificilmente as extremidades seriam preservadas.
Outros que não são recuperados como fósseis são aqueles cujas paredes são impregnadas de sílica ou de carbonato de cálcio, pois estas substâncias seriam dissolvidas na maceração química que caracteriza uma das etapas da preparação palinológica. Caso se pretenda estudar esses tipos de tricomas ricos em cálcio e sílica, deve-se recorrer a uma preparação diferente da palinológica.
TIPOLOGIA DOS TRICOMAS FÔSSEIS ENCONTRADOS
Para o estudo dos morfotipos fósseis, adotou-se uma classificação baseada na literatura, procurando utilizar os termos já existentes, mas adaptados aos tipos encontrados.
Foi elaborado um esquema de classificação adaptado a partir das propostas de Theobald et al. (1979), Llamas et al. (1995) e Webster et al. (1996). A classificação de Payne (1978) não foi utilizada, por incluir critérios funcionais que não são aplicáveis a tricomas fósseis.
Foram adotados 12 morfotipos básicos: simples, em forma T, em forma Y (com variantes U, V e J), duplo, triplo, fasciculado, fasciculado tufoso, multirradiado, multirradiado tufoso, estrelado tridimensional, estrelado tridimensional tufoso e estrelado bidimensional ou planar (Figura 2).
A natureza tufosa não foi considerada nos cinco primeiros morfotipos básicos (simples a triplo), por não ser característica preponderante em tricomas com poucas ramificações, mas pode ser usada para a caracterização de seus submorfotipos.
DESCRIÇÃO DE PARÂMETROS MORFOLÓGICOS
Para sistematizar a definição de morfotipos e seus subtipos, foram considerados os seguintes parâmetros morfológicos: número de ramificações ou raios, diâmetro relativo, curvatura, número de células, tipo da base, forma da extremidade, ângulo entre as ramificações ou raios, forma do centro (em tricomas estrelados) e pedicelo.
Número de ramificações ou raios: o número de ramificações pode variar de um, nos tricomas simples, a dezenas, nos multirradiados e estrelados. As ramificações são também chamadas de 'raios' no caso de tricomas multirradiados abertos e estrelados.
Diâmetro relativo: quanto ao diâmetro relativo, os tricomas podem ser classificados em delgados (razão comprimento/diâmetro médio ≥ 10) e robustos (razão comprimento/diâmetro médio < 10).
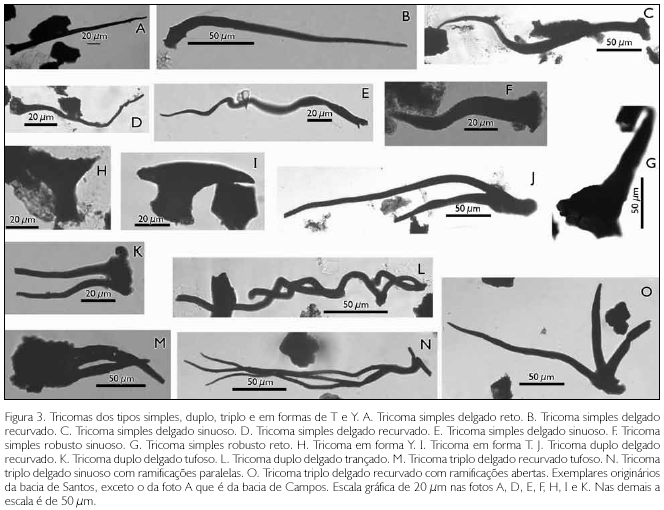
Curvatura: em termos de curvatura, os tricomas podem ser classificados em retos (sem curvatura) (Figura 3A), recurvados (apenas uma curva) (Figura 3 B) ou sinuosos (duas ou mais curvas) (Figuras 3C, 3E e 3F).
Número de células: pode ser unicelular ou multicelular, dependendo do número de células em cada ramificação ou raio. Em materiais botânicos não fossilizados, a distinção é fácil, já que as ramificações multicelulares apresentam um padrão de segmentação que lembra o de um caule de bambu. No entanto, a distinção não é tão evidente no material fóssil, que se apresenta opaco em função da carbonificação e/ou carbonização. Nesse caso, o caráter multicelular seria notável somente se a segmentação se manifestar no contorno externo.
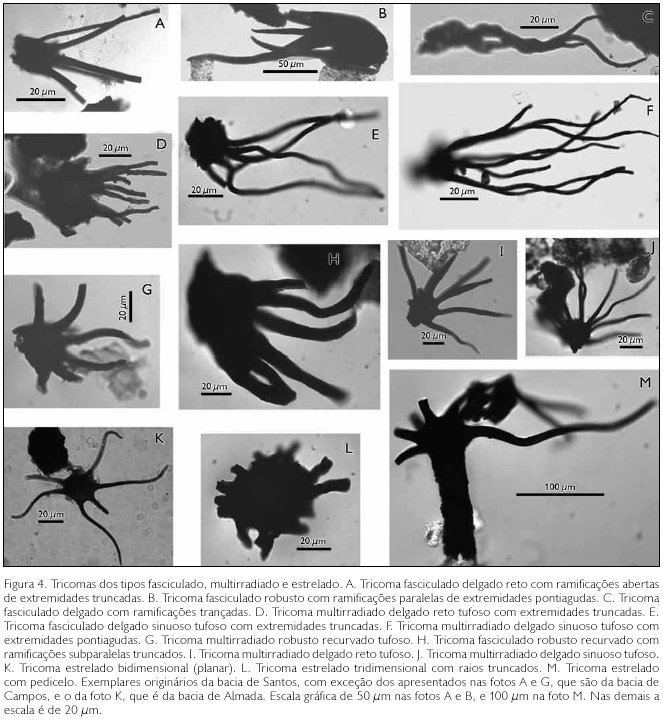
Forma da extremidade: pode ser pontiaguda, arredondada, capitada ou truncada. Aqui, cabe uma discussão quanto à validade da extremidade truncada como uma das características para descrição morfológica. Extremidades truncadas, por si só, não podem constituir um critério morfotaxonômico, mas, se todas as extremidades estiverem aparadas mais ou menos na mesma altura, isto poderá constituir uma característica morfológica importante (Figuras 4A, 4H e 4L). Extremidades truncadas de modo sistemático e uniforme podem indicar a ocorrência de tricomas glandulares, cujas terminações ou extremidades não se preservaram no material fóssil. Uma forma desse tipo foi registrada por Krings et al. (2003) numa pteridófita fóssil do Carbonífero (Paleozóico), cujas extremidades dos tricomas seriam "explosivas", à semelhança dos tricomas glandulares de Sicana odorifera (Vell.) Naudin (Cucurbitaceae). A outra possibilidade é que a perda das extremidades seja decorrente da natureza histoquímica, como no caso do estudo de Milanez & Machado (2008), que demonstraram que as extremidades dos tricomas de Microlepis oleaefolia (Melastomataceae) são de composição péctica, de difícil preservação. Nesse caso, a possibilidade de fossilização das extremidades seria praticamente nula.
Tipo da base: considera-se normal, caso não apresente uma diferenciação na base; expandida, quando a base é visivelmente maior do que o diâmetro do restante do tricoma (Figura 5A, 5B e 5C), e tufosa, quando apresenta um aglomerado de células (Figuras 5D, 5E e 5F).

Ângulo entre as ramificações: o ângulo entre as ramificações ou raios constitui um critério importante para distinguir vários morfotipos. Por exemplo, é com base no ângulo que se distinguem os tricomas dos tipos duplo, em forma T e em forma Y Para o presente estudo, convencionou-se adotar as medidas de ângulo entre os raios extremos, para distinguir os tipos fasciculado (< 90o), multirradiado (90o - 200o) e estrelado tridimensional (200o - 360o). Entre os fasciculados, estabeleceu-se a seguinte classificação em termos de abertura angular: paralelo (< 5o), subparalelo (5o - 30o) e aberto (> 30o).
Forma da parte central: em tricomas estrelados, a feição da parte central pode constituir critérios para o estabelecimento de subtipos. Existem basicamente três morfotipos da parte central: tufoso, maciço e de contato simples (contato direto entre as ramificações, sem a intervenção da estrutura central).
Presença de pedicelo: teoricamente, pedicelos podem estar presentes tanto nos tricomas fasciculados como nos estrelados, mas, neste estudo, foi encontrado apenas um exemplar de tricoma estrelado pedicelado (Figura 4M).
DESCRIÇÃO DOS TIPOS E SUBTIPOS
Seguem as descrições de cada morfotipo e respectivos subtipos. Convém lembrar que nem todos os subtipos foram efetivamente registrados no material estudado. No entanto, eles foram listados por serem tipos constituídos pelas combinações viáveis de características morfológicas, fato comprovado pela existência de formas equivalentes em plantas vivas. Este procedimento permitirá a incorporação de novos morfotipos ao banco de dados paleontológicos, logo que forem encontrados.
1. Tipo simples: tipo dotado de uma única ramificação; chamado frequentemente de "pelo" (hair) na literatura (e.g., Ayensu, 1974; Andrzejewska-Golec, 1998, 2000, 2003; Abdel-Khalik, 2005). Para os subtipos mencionados abaixo podem ser aplicados ainda os seguintes atributos: número de células (unicelular ou multicelular); tipo da base (normal ou expandida); forma da extremidade (pontiaguda, arredondada, capitada ou truncada) e curvatura.
a. Subtipo simples delgado reto (Figura 3A)
b. Subtipo simples delgado recurvado (Figuras 3B, 3D e 5B)
c. Subtipo simples delgado sinuoso (Figuras 3C e 3E)
d. Subtipo simples robusto reto (Figura 3G)
e. Subtipo simples robusto recurvado
f. Subtipo simples robusto sinuoso (Figura 3F)
2. Tipo em forma T: tipo dotado de duas ramificações que, juntamente com a base, constitui uma forma semelhante à letra T (Figura 3I).
3. Tipo em forma Y: tipo dotado de duas ramificações que, juntamente com a base, constitui uma forma semelhante à letra Y (Figura 3 H). Esta denominação é utilizada por diversos autores (e.g., Abdel-Khalik, 2005; Theobald et al., 1979). Dependendo da curvatura das ramificações e do grau de simetria, este tipo admite as formas variantes U, V e J (Theobald et al., 1979).
a. Tipo em forma Y (Figura 3H)
b. Tipo em forma U
c. Tipo em forma V
d. Tipo em forma J
4. Tipo duplo: tipo dotado de duas ramificações orientadas mais ou menos na mesma direção; na literatura, aparece também como "twin hairs" (Sancho & Katinas, 2002) e seria um caso particular de "two-armed trichome" (Theobald et al., 1979). Para os subtipos mencionados abaixo, podem ser aplicados ainda os seguintes atributos: número de células (unicelular ou multicelular); tipo da base (normal, expandida ou tufosa); forma da extremidade (pontiaguda, arredondada, capitada ou truncada); ângulo entre as ramificações (paralelo, subparalelo ou aberto) e curvatura.
a. Subtipo duplo delgado reto
b. Subtipo duplo delgado recurvado (Figuras 3J e 3K)
c. Subtipo duplo delgado sinuoso
d. Subtipo duplo delgado trançado (Figura 3L)
e. Subtipo duplo robusto reto
f. Subtipo duplo robusto recurvado
g. Subtipo duplo robusto sinuoso
5. Tipo triplo: tipo dotado de três ramificações mais ou menos subparalelas; na literatura, aparece também como "three-armed trichome" (Theobald et al., 1979). Para os subtipos mencionados abaixo, podem ser aplicados os seguintes atributos: número de células (unicelular ou multicelular); tipo da base (normal, expandida ou tufosa); forma da extremidade (pontiaguda, arredondada, capitada ou truncada); ângulo entre as ramificações (paralelo, subparalelo ou aberto) e curvatura.
a. Subtipo triplo delgado reto
b. Subtipo triplo delgado recurvado (Figuras 3M e 3O)
c. Subtipo triplo delgado sinuoso (Figura 3N)
d. Subtipo triplo delgado trançado
e. Subtipo triplo robusto reto
f. Subtipo triplo robusto recurvado
g. Subtipo triplo robusto sinuoso
6. Tipo fasciculado: tipo com ramificações reunidas em forma de feixe; o ângulo formado entre as ramificações extremas deve ser menor do que 90o; tipicamente, seria dotado de quatro a sete ramificações, segundo o critério estabelecido a partir da combinação das definições de Llamas et al. (1995) e Webster et al. (1996), mas, neste trabalho, adotou-se o ângulo como um critério preponderante. Este tipo equivale, em parte, aos seguintes morfotipos mencionados na literatura: stellate-multiangulate (Theobald et al., 1979), fasciculate (Llamas et al., 1995; Webster et al., 1996), multiradiate (Llamas et al., 1995). Para os subtipos mencionados abaixo, podem ser aplicados os seguintes atributos: número de células em cada ramificação (unicelular ou multicelular); forma da extremidade (pontiaguda, arredondada, capitada ou truncada); ângulo entre as ramificações (paralelo, subparalelo ou aberto) e curvatura.
a. Subtipo fasciculado delgado reto (Figura 4A)
b. Subtipo fasciculado delgado recurvado
c. Subtipo fasciculado delgado sinuoso (Figura 5C)
d. Subtipo fasciculado delgado trançado (Figura 4C)
e. Subtipo fasciculado robusto reto
f. Subtipo fasciculado robusto recurvado (Figura 4B)
g. Subtipo fasciculado robusto sinuoso
7. Tipo fasciculado tufoso: possui as mesmas características do morfotipo anterior, diferindo apenas pela presença de base tufosa.
a. Subtipo fasciculado delgado reto tufoso
b. Subtipo fasciculado delgado recurvado tufoso
c. Subtipo fasciculado delgado sinuoso tufoso (Figuras 4E e 5D)
d. Subtipo fasciculado delgado trançado tufoso
e. Subtipo fasciculado robusto reto tufoso
f. Subtipo fasciculado robusto recurvado tufoso (Figura 4H)
g. Subtipo fasciculado robusto sinuoso tufoso
8. Tipo multirradiado: tipo com ramificações (raios) reunidas em forma de feixe; o ângulo formado pelas ramificações extremas varia de 90o a 200o; dotado de oito ou mais raios, critério este adotado a partir da definição de Webster et al. (1996); equivale exatamente ao morfotipo multiradiate, de Webster et al. (1996), e, em parte, ao stellate-multiangulate, de Theobald et al. (1979), e multiradiate, de Llamas et al. (1995). Para os subtipos mencionados abaixo, podem ser aplicados ainda os seguintes atributos: número de células em cada raio (unicelular ou multicelular); forma da extremidade (pontiaguda, arredondada, capitada ou truncada); ângulo entre as ramificações e curvatura.
a. Subtipo multirradiado delgado reto
b. Subtipo multirradiado delgado recurvado
c. Subtipo multirradiado delgado sinuoso (Figura 4F)
d. Subtipo multirradiado robusto reto
e. Subtipo multirradiado robusto recurvado
f. Subtipo multirradiado robusto sinuoso
9. Tipo multirradiado tufoso: possui as mesmas características do morfotipo anterior, diferindo apenas pela presença de base tufosa.
a. Subtipo multirradiado delgado reto tufoso (Figuras 4D e 4I)
b. Subtipo multirradiado delgado recurvado tufoso
c. Subtipo multirradiado delgado sinuoso tufoso (Figura 4J)
d. Subtipo multirradiado robusto reto tufoso
e. Subtipo multirradiado robusto recurvado tufoso (Figura 4G)
f. Subtipo multirradiado robusto sinuoso tufoso
10. Tipo estrelado tridimensional: tipo com ramificações (raios) reunidas em forma de estrela; dotado de oito ou mais raios que se projetam em quase todas as direções (o ângulo formado pelas ramificações extremas mede 200o, no mínimo); equivale exatamente ao morfotipo stellate, de Webster et al. (1996), e, em parte, ao stellate-multiangulate, de Theobald et al. (1979), e stellate e fused-stellate, de Llamas et al. (1995). Para os subtipos mencionados abaixo, podem ser aplicados os seguintes atributos: número de células em cada raio (unicelular ou multicelular); forma da extremidade (pontiaguda, arredondada, capitada ou truncada); forma da parte central; presença de pedicelo e curvatura.
a. Subtipo estrelado tridimensional delgado reto
b. Subtipo estrelado tridimensional delgado recurvado
c. Subtipo estrelado tridimensional delgado sinuoso
d. Subtipo estrelado tridimensional robusto reto
e. Subtipo estrelado tridimensional robusto recurvado
f. Subtipo estrelado tridimensional robusto sinuoso
11. Tipo estrelado tridimensional tufoso: possui as mesmas características do morfotipo anterior, diferindo apenas pela presença de base tufosa.
a. Subtipo estrelado tridimensional delgado reto tufoso
b. Subtipo estrelado tridimensional delgado recurvado tufoso
c. Subtipo estrelado tridimensional delgado sinuoso tufoso
d. Subtipo estrelado tridimensional robusto reto tufoso
e. Subtipo estrelado tridimensional robusto recurvado tufoso
f. Subtipo estrelado tridimensional robusto sinuoso tufoso
12.Tipo estrelado bidimensional (planar): tipo com cinco ou mais raios, reunidos em forma de estrela, que se projetam em todas as direções num único plano; equivale exatamente ao morfotipo stellate-rotate, de Theobald et al. (1979), e aos morfotipos stellate-rotate, stellate-lepidote e dentate-lepidote, de Webster et al. (1996), e, em parte, ao fused-stellate, de Llamas et al. (1995). Potencialmente, podem ser aplicados os seguintes atributos para estabelecer seus subtipos: número de células em cada raio (unicelular ou multicelular); feição da ponta (pontiaguda, arredondada, capitada ou truncada); natureza da junção dos raios (tufoso ou não tufoso); comprimento relativo dos raios; e proporção da área fundida. No entanto, no presente estudo, foi encontrado apenas um exemplar deste tipo, proveniente do Maastrichtiano da bacia de Almada, que pode ser classificado como 'tipo estrelado planar unicelular não-tufoso com raio delgado com extremidade pontiaguda e área fundida relativamente pequena' (Figura 4K).
CONCLUSÃO
A proposta de classificação estabelecida neste trabalho visa sistematizar o registro de tricomas fósseis recuperados em preparações paleopalinológicas de amostras oriundas do Cretáceo Superior. A sistematização do registro é importante para consolidar a validade de tricomas como fósseis-guias utilizáveis, o que deve ser alcançado com o acúmulo de dados de sua ocorrência. Mas, mesmo antes de atingir a massa crítica de dados, pode-se dizer que muitos dos morfotipos aqui registrados devem mesmo servir de ferramenta bioestratigráfica, já que são desconhecidos em outros níveis estratigráficos, devendo, portanto, ser exclusivos da seção analisada. Um exemplo típico é o único tricoma estrelado planar, que é do Maastrichtiano, e que não foi encontrado no Santoniano.
A viabilização do uso estratigráfico de tricomas torna-se especialmente importante no Santoniano das bacias de Campos e Santos, onde, em consequência dos paleoincêndios ocorridos na época, há escassez de palinomorfos stricto sensu (Arai et al., 2006; Arai, 2008). Enfim, os tricomas fósseis devem ser submetidos à mesma linha de pesquisa já iniciada para o estudo de esclerócitos (esclereídes fósseis) por Duarte & Arai (2008, 2010).
Outra contribuição que os tricomas podem trazer, caso seja possível o estabelecimento da relação entre os morfotipos e a família botânica das plantas genitoras, é a informação relativa à paleoflora e ao paleoambiente, uma vez que eles podem fornecer informações paleoambientais de diversas maneiras. Metcalfe & Chalk (1950) notaram que o comprimento, o tamanho e a densidade de tricomas são muito mais variáveis em resposta a condições ambientais do que a variação de tipos de tricomas em um determinado táxon. Por outro lado, frequentemente a presença de um tipo particular pode colaborar para a determinação de categorias taxonômicas vegetais, como espécie, gênero e família.
Outro aspecto que deve ser lembrado é a familiarização dos paleopalinólogos com tricomas. Na falta de informação, alguns pesquisadores podem até cometer identificações equivocadas, como ocorre com os exemplares de tricomas robustos mostrados nas Figuras 3I e 3G, que poderiam ser classificados como escolecodontes por olhos mais desavisados. Outra confusão que ocorre é a identificação equivocada de tricomas estrelados bidimensionais (planares) como sendo corpos frutíferos de fungos da família Microthyriales (e.g., estampa 15, figura 23 da tese de doutorado de H. S. El Atfy, defendida em 2008, intitulada "Applied palynological studies on the Cretaceus sucession of the GPTSW-7 well, Abu Sennan Concession, Western Desert, Egypt").
Sendo assim, estudos mais aprofundados destas estruturas lignificadas, tratadas no presente trabalho como palinomorfos lato sensu, reconhecidas e classificadas como tricomas, são de muita importância e devem ter um enfoque relevante nos futuros estudos palinológicos.
AGRADECIMENTOS
Os autores desejam manifestar sua gratidão ao Prof. Dr. Ricardo Cardoso Vieira (Universidade Federal do Rio de Janeiro) e ao Prof. Arno Fritz das Neves Brandes (Instituto de Pesquisa, Jardim Botânico do Rio de Janeiro), pelo empréstimo do material bibliográfico e sugestões oportunas; ao Dr. José Ricardo Maizatto (Centro de Pesquisas e Desenvolvimento Leopoldo Américo Miguez de Mello, Petrobras), pela leitura minuciosa do manuscrito; ao Prof. Geoffrey Playford (Universidade de Queensland, Brisbane, Austrália), pela revisão do abstract; e à Petrobras, pela autorização da publicação deste trabalho.
REFERÊNCIAS
ABDEL-KHALIK, K., 2005. Morphological studies on trichomes of Brassicaceae in Egypt and taxonomic significance. Acta Botânica Croatica 64(1): 57-73.
ALQUINI, Y., C. BONA, M. R. T. BOEGER, C. G. COSTA & C. F. BARROS, 2003. Epiderme. In: B. APPEZZATO-DA-GLÓRIA & S. M. CARMELLO-GUERREIRO (Eds.): Anatomia Vegetal: (1): 87-97. Editora UFV, Viçosa.
ANDERSON, D. B. & T. KERR, 1938. Growth and structure of cotton fiber. Industry and Engineering Chemistry 30: 48-54.
ANDRZEJEWSKA-GOLEC, E., 1998. Microhairs of Littorella uniflora (L.) Asch. (Plantaginaceae). Feddes Repertorium 109(7-8): 521-526.
ANDRZEJEWSKA-GOLEC, E., 2000. The hairs in species of Plantago L. section Palaeopsyllium Pilg. (Plantaginaceae). Feddes Repertorium 111(1-2): 9-14.
ANDRZEJEWSKA-GOLEC, E., 2003. The hairs of Plantago reniformis Beck, section Eremopsyllium Pilg. (Plantaginaceae). Feddes Repertorium 114(3-4): 204-207.
ARAI, M., 2008. Evidências palinológicas de vulcanismo incendiário no Cretáceo Superior das bacias do Sudeste Brasileiro. Anais do 4o Simpósio de Vulcanismo e Ambientes Associados. Disponível em: <http://jasper.rc.unesp.br/vulcanismoArabalhos/arai.pdf>. Acesso em: 5 agosto 2009.
ARAI, M., C. C. LANA, C. V. ARAÚJO & T. R. MENEZES, 2006. O grande incêndio do Santoniano: considerações geológicas e exploratórias. Boletim de Geociências da Petrobras 14(1): 125-131.
AYENSU, E. S., 1974. Leaf anatomy and systematics of New World Velloziaceae. Smithsonian Contributions to Botany 15: 1-125.
BEYRICH, H., 1943. Über die Membranverkieselung einiger Pflanzenhaare. Flora 36: 313-324.
CORREIA, G. A., J. R. C. MENEZES, G. V. BUENO & E. J. J. MARQUES, 2005. Identificação de uma estrutura de impacto no Cretáceo Superior da Bacia de Santos em sísmica de reflexão 3D. Boletim de Geociências da Petrobras 13(1): 123-127.
DUARTE, S. G. & M. ARAI, 2008. Esclerócitos: estudo comparativo entre os fósseis e os de plantas viventes. Boletim de Resumos do Simpósio Brasileiro de Paleobotânica e Palinologia 12: 57.
DUARTE, S. G. & M. ARAI, 2010. Esclerócitos: proposta de uma nova categoria de palinomorfos lato sensu. Revista Brasileira de Paleontologia 13(1): 67-76.
ESAU, K., 1953. Plant Anatomy: 1-735. John Wiley & Sons, Inc., New York.
FAHN, A., 1986. Structure and functional properties oftrichomes of xeromorphic leaves. Annales of Botany 57: 631-637.
FAHN, A., 1990. Plant Anatomy: 1-588. Pergamon Press, New York.
FAHN, A. & D. I. CUTLER, 1992. Xerophytes: 1-176. Gebrüder Broentaeger, Berlin.
FREIRE, S. E., E. URTUBEY & D. A. GIULIANO, 2007. Epidermal characters of Baccharis (Asteraceae) species used in traditional medicine. Caldasia 29(1): 23-38.
KRINGS, M., D. W. KELLOGG, H. KERP & T. N. TAYLOR, 2003. Trichomes of the seed fern Blanzyopteris praedentata: implications for plant-insect interaction in the Late Carboniferous. Botanical Journal of Linnean Society 141: 133-149.
LLAMAS, F., C. PEREZ-MORALES, C. ACEDO & A. PENAS, 1995. Foliar trichomes of the evergreen and semideciduous species of the genus Quercus (Fagaceae) in the Iberian Peninsula. Botanical Journal of the Linnean Society 117: 47-57.
LODARI, C., K. HATTORI & Y. FUTSUHARA, 1989. Morphological difference on leaf surface and pollen grains in genus Artemisia. Japanese Journal of Breeding 39: 9-14.
METCALFE, C. R., 1960. Anatomy of the Monocotyledons. I. Gramineae: 1-731. Clarendon Press, Oxford.
METCALFE, C. R. & L. CHALK, 1950. Anatomy of the Dicotyledons. II : 1-1500. Clarendon Press, Oxford.
METCALFE, C. R. & L. CHALK, 1988. Anatomy of the Dicotyledons. I, 2nd edition: 1-276. Clarendon Press, Oxford.
MILANEZ, C. R. D. & S. R. MACHADO, 2008. Leaf emergences in Microlepis oleaefolia (D.C.) Triana (Melastomataceae) and their probable function: an anatomical and ultrastructural study. Micron 39(7): 884-890.
NETOLITZKY, F., 1932. Die Pflanzenhaare. In: K. LINSBAUER (Ed.): Handbuch der Pflanzenatomie: 4: 20-23. Gebruder Borntraeger, Berlin.
PAYNE, W. W., 1978. A glossary of plant hair terminology. Brittonia 30: 239-255.
SANCHO, G. & L. KATINAS, 2002. Are the trichomes in corollas of Mutisieae (Asteraceae) really twin hairs? Botanical Journal of the Linnean Society 140: 427-433.
THEOBALD, W. L., J. L. KRAHULIK & R. C. ROLLINS, 1979. Trichome description and classification. In: C. R. METCALFE & L. CHALK (Eds.): Anatomy of the Dicotyledons. I: 2nd edition: 40-53. Clarendon Press, Oxford.
TOMLINSON, P. B., 1969. Commelinales-Zingiberales. In: C. R. METCALFE (Ed.): Anatomy of the Monocotyledons: 193-294. Clarendon Press, Oxford.
UESUGUI, N., 1979. Palinologia: técnicas de tratamento de amostras. Boletim Técnico da Petrobras 22(4): 229-240.
WEBSTER, G. L., M. J. DEL-ARCO-AGUILAR & B. A. SMITH, 1996. Systematic distribution of foliar trichome types in Croton (Euphorbiaceae). Botanical Journal of the Linnean Society 121: 41-57.
ZALÁN, P. V. & J. A. B. OLIVEIRA, 2005. Origem e evolução estrutural do sistema de riftes cenozóicos do Sudeste do Brasil. Boletim de Geociências da Petrobras 13(2): 269-300.
 Endereço para correspondência:
Endereço para correspondência:
Museu Paraense Emílio Goeldi
Editor do Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais
Av. Magalhães Barata, 376
São Braz – CEP 66040-170
Belém - PA - Brasil
Caixa Postal 399
Telefone: 55-91-3182-3246
Fax: 55-91-3249-6373
E-mail:boletim.naturais@museu-goeldi.br
Recebido: 15/01/2010
Aprovado: 06/08/2010