










Serviços Personalizados
Journal

Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Pan-Amazônica de Saúde
versão impressa ISSN 2176-6215versão On-line ISSN 2176-6223
Rev Pan-Amaz Saude v.1 n.2 Ananindeua jun. 2010
http://dx.doi.org/10.5123/S2176-62232010000200012
ARTIGO ORIGINAL | ORIGINAL ARTICLE | ARTÍCULO ORIGINAL
Partial genetic characterization of Seoul hantavirus in rats from Buenos Aires City, Argentina, and generation of a Seoul recombinant nucleoprotein antigen
Caracterização genética parcial do hantavírus Seoul em ratazanas provenientes de Buenos Aires, Argentina, e geração de um antígeno a partir da nucleoproteína recombinante do vírus Seoul
Caracterización genética parcial del hantavirus Seoul en ratas provenientes de Buenos Aires, Argentina, y generación de un antígeno a partir de la nucleoproteína recombinante del virus Seoul
Paula Julieta PadulaI; Valeria Paula MartínezI; Gerardo Rubén CuetoII; Regino CaviaII; Olga Virginia SuárezII
IDepartamento de Virología, Instituto Nacional de Enfermedades Infecciosas, Administración Nacional de Laboratorios e Institutos de Salud "Dr. Carlos G. Malbrán", Buenos Aires, Argentina
IILaboratorio de Ecología de Roedores, Departamento de Ecología Genética y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires, Argentina
Endereço para correspondência
Correspondence
Dirección para correspondencia
ABSTRACT
Hemorrhagic fever with renal syndrome (HFRS) is a severe infectious disease characterized by fever, hemorrhage, renal impairment, and thrombocytopenia. At least seven hantaviruses cause HFRS: Hantaan, Seoul (SEOV) (distributed worldwide), Dobrava/Belgrade, Saaremaa, Amur, Thailand and Puumala. To investigate the epidemiology of HFRS and virus transmission in Argentina, we constructed a prokaryotic plasmid encoding the SEOV rNP, of 430 amino acids. After expression, the rNP was tested as an antigen for use in an enzyme-linked immunosorbent assay for infection diagnosis. To determine the current level of virus transmission in wild brown rats or Norway rats (Rattus norvegicus) captured in Buenos Aires City, Argentina, we tested tissues from rats that were determined to be serologically positive for the SEOV, and the viral genome were detected by RT-PCR using specific primers for two fragments of M segment-encoding Gn and Gc proteins. The viral genome was detected in 11 of 21 seropositive rats (52.4%) captured in two parklands. Sequence analysis of a 333-nt region of the Gc-encoding M segment revealed 97% and 96% identity with strains of SEOV from Baltimore and Brazil, respectively. Our genetic data confirm a very low diversity among SEOV virus strains.
Keywords: Hantavirus; Seoul virus; Rats; Prokaryotic Cells; Recombinant Proteins; Enzyme-Linked Immunosorbent Assay.
RESUMO
A febre hemorrágica com síndrome renal (FHSR) é uma doença grave, caracterizada por febre, hemorragia, falência renal e trombocitopenia. Pelo menos sete hantavírus causam a FHSR: Hantaan, Seoul (SEOV) (de distribuição global), Dobrava-Belgrade, Saaremaa, Amur, Thailand e Puumala. Para investigar a epidemiologia da FHRS e a transmissão viral na Argentina, criamos um plasmídio procariótico que "codifica" a nucleoproteína recombinante do vírus SEOV de 430 aminoácidos. Após a expressão, a nucleoproteína recombinante foi testada como antígeno para uso em ensaio imunoenzimático (ELISA) para diagnóstico da infecção. Para determinar o nível atual de transmissão viral em populações de ratos-marrons ou ratazanas (Rattus norvegicus) capturadas na cidade de Buenos Aires, Argentina, testamos tecidos de ratos selecionados para serem sorologicamente positivos para o vírus SEOV, e o seu genoma viral foi detectado após submetido a RT-PCR utilizando primers específicos para dois fragmentos de proteínas Gn e Gc codificadas pelo segmento M. O genoma viral foi detectado em 11 das 21 ratazanas soropositivas (52,4%), previamente capturadas em dois parques. A análise sequencial de uma região gênica (333 nt) do segmento M "codificador" da proteína Gc apresentou 97% e 96% de similaridade com as cepas de SEOV coletadas em Baltimore e no Brasil, respectivamente. Os dados genéticos levantados confirmam a informação de que há uma diversidade muito pequena entre as cepas do vírus SEOV.
Palavras-chave: Hantavirus; Vírus Seul; Ratos; Células Procarióticas; Proteínas Recombinantes; ELISA.
RESUMEN
La fiebre hemorrágica con síndrome renal (FHSR) es una enfermedad grave, caracterizada por fiebre, hemorragia, falencia renal y trombocitopenia. Al menos siete hantavirus causan la FHSR: Hantaan, Seoul (SEOV) (de distribución global), Dobrava-Belgrade, Saaremaa, Amur, Thailand y Puumala. Para investigar la epidemiología de la FHRS y la transmisión viral en Argentina, creamos un plásmido procariotas que "codifica" la nucleoproteína recombinante del virus SEOV de 430 aminoácidos. Luego de la expresión, la nucleoproteína recombinante fue probada como antígeno para uso en ensayo inmunoenzimático (ELISA) para diagnóstico de la infección. Para determinar el nivel actual de transmisión viral en poblaciones de ratas marrones o ratas (Rattus norvegicus) capturadas en la ciudad de Buenos Aires, Argentina, analizamos tejidos de ratas seleccionadas para ser serológicamente positivas para el virus SEOV, y su genoma viral fue detectado luego de sometido a RT-PCR utilizando primers específicos para dos fragmentos de proteínas Gn y Gc codificadas por el segmento M. El genoma viral fue detectado en 11 de las 21 ratas seropositivas (52,4%), previamente capturadas en dos parques. El análisis secuencial de una región génica (333 nt) del segmento M "codificador" de la proteína Gc presentó un 97% y un 96% de similitud con las cepas de SEOV colectadas en Baltimore y en Brasil, respectivamente. Los datos genéticos listados confirman la información de que hay una diversidad muy pequeña entre las cepas del virus SEOV.
Palabras clave: Hantavirus; Virus Seoul; Ratas; Células Procarióticas; Proteínas Recombinantes; Prueba ELISA.
INTRODUCTION
Hantaviruses (family Bunyaviridae, genus Hantavirus) are rodent-borne viruses that cause two serious diseases. In Eurasia, hemorrhagic fever with renal syndrome (HFRS) is a severe infectious disease caused by at least four antigenically and genetically distinct hantaviruses defined as different serotypes: Hantaan (HTNV), Seoul (SEOV), Dobrava/Belgrade (DOBV), and Puumala (PUUV)21. In the Americas, sigmodontine-associated hantaviruses, such as Sin Nombre virus (SNV) in North America and Andes virus (ANDV) in South America, cause hantavirus pulmonary syndrome17,14. The hantaviruses have a tripartite, negative-stranded RNA genome that encodes an RNA-dependent RNA polymerase, two envelope glycoproteins (Gn and Gc), and a nucleocapsid protein (NP)20.
In nature, each hantavirus is predominantly carried by rodents of separate species, and diseases caused by infections with these viruses are confined to the region corresponding to the distribution of the rodent host22. However, as Rattus species are cosmopolitan, SEOV has the potential to cause human disease worldwide due to the distribution of its infected hosts, black rats (R. rattus) and Norway rats (R. notvegicus)13, through international freight transportation. At present, there are only a few reports of confirmed human SEOV infections outside Asia, including the USA7 and Brazil9. To date, HFRS resulting from SEOV infection has been confined mainly to Asian countries19. In Buenos Aires, SEOV has been serologically detected in Norway rats since 198512,23,26; however, no HFRS cases caused by SEOV have been registered23. Recently, a seroprevalence study of SEOV infection in Norway rats from Buenos Aires City revealed a positivity of 11.9%2, but the viral genome was not characterized.
Due to the hazardous nature of hantaviruses, their slow replication, and low and variable yield in cell culture, recombinant hantavirus proteins have been produced for use as antigens in serological assays including those for SEOV10,27,25,5,4,18,1.
Here, we present the first genetic evidence of SEOV in Norway rats in Buenos Aires City, Argentina. We also generated a SEOV recombinant nucleocapsid protein (rNP) antigen for use in serologic tests.
MATERIALS AND METHODS
WILD-CAUGHT ANIMALS
From 2003 to 2005, wild rats were trapped from several locations in Buenos Aires City, Argentina, such as urban parklands larger than 20 hectares, urban residential areas with high densities of humans and shantytowns. During the study, 151 Norway rats and 33 black rats were captured, with a total trapping effort of 14,088 trap-nights2.
Blood samples were collected by cardiac puncture from all individuals captured and were used for serological analysis. The lung, liver and kidney tissues were removed from each rodent and stored in liquid nitrogen until tested.
CLONING, EXPRESSION AND PURIFICATION OF SEOV EXPRESSING rNP
For cloning, we used the pGEM-T vector and DH5α Escherichia coli strain grown in Luria-Bertani medium. The SEOV Sapporo rat strain stock virus (kindly provided by Jay Hooper and Connie Schmaljohn, United States Army Medical Research Institute of Infectious Diseases, Fort Detrick, Frederick, MD) was amplified in Vero cells, and the viral RNA was extracted from the cell supernatant. To generate a plasmid containing the complete ORF of the N protein, we conducted RT-PCR with primers designated to contain restriction sites annealing immediately upstream of the N start codon and immediately down-stream of the N stop codon. The pGEM-T ligated plasmid with the cDNA insert was transformed into DH5α E. coli.
For recombinant protein expression, we used pRSET B (Invitrogen Life Technologies) to transform the plasmid into the BL21 (DE3) E. coli strain, according to the manufacturer's instructions, and selected for ampicillin-resistant transformants. After cell lysis and centrifugation, the supernatant was purified by Ni-affinity chromatography. The purified proteins were analyzed by glycine, sodium dodecyl sulfate, 10% polyacrylamide gel electrophoresis (SDS-PAGE).
ENZYME-LINKED IMMUNOSORBENT ASSAY
An enzyme-linked immunosorbent assay (ELISA) was performed on whole blood samples for detection of IgG antibodies, as previously described for ANDV18. Briefly, plates were coated overnight at 4° C with SEOV rNP and control recombinant antigen or Vero E6 cells infected with SEOV and uninfected Vero E6 cells. Samples were diluted 1:200 in phosphate-buffered saline-Tween 20 and added in duplicate to antigen-coated wells containing infected or uninfected Vero E6 cells. The plates were washed with the same buffer and incubated at 37° C for 1 h with secondary antibody, alkaline phosphatase-conjugated anti-rat IgG (H + L; Kirkegaard and Perry Laboratories, Gaithersburg, MD). The plates were incubated for one hour at 37° C, and the substrate for peroxidase was added. The optical density (OD) was measured at 405 nm, and the average OD for each set of uninfected Vero E6 duplicates or control recombinant antigen was subtracted from the average OD for each set of infected Vero E6 duplicates or SEOV rNP antigen, respectively. Samples were considered positive if the difference between OD values was greater than 0.3. Because sigmodontine-borne hantaviruses have been reported to be circulating in Buenos Aires and its surrounding areas, we also analyzed rat sera with low SEOV-ELISA test values using ANDV recombinant antigen, as previously described18.
The end-point ELISA titer was defined as the reciprocal of the highest dilution at which the OD value was 0.3 or greater.
ISOLATION OF RNA, AMPLIFICATION, SEQUENCING AND DNA ANALYSIS
Total RNA, extracted from lung or kidney tissues of rodents with Trizol reagent (Invitrogen Life Technologies®), was reverse transcribed using AMV (Promega Corp.®) and specific primers following the instructions of the manufacturer. Hemi-nested and single round RT-PCR procedures were performed to amplify two partial M-segment hantaviral RNA fragments. To amplify Gn fragment of 437 nt (6 to 442), we used primers P0 (5'- TAGTAGTAGACTCCGC-3') with P10 (5' - GTTTGATTACAGGCCAGATCATAACA-3') for the first round and P57 (5'-GACTCCGCAAGAAGAAGC-3') with P10 for the second round. A sequence of 385 nt was obtained. For the analysis of Gc fragment (333 nt from 1984 to 2316) were sequenced24. All positions were related to the SEOV Sapporo strain. The PCR products were separated by agarose gel electrophoresis, stained with ethidium bromide, and visualized under UV light. The products were purified and sequenced directly with an automated DNA sequencer (ABI PRISM® 3100-Avant Genetic Analyzer System, Perkin-Elmer, USA).
The Genbank accession numbers of the previously published reference strains used are as follows: AF324901 (ANDV); AF00578 Laguna Negra virus (LNV); L25783 (SNV); M29979 (PUUV); NC005228 (TULV); L33685 (DOBV); M14627 (HTNV); SEOV strain U00151 (Baltimore), SEOV strain U00460 (Brazil), SEOV Chinese strains: DQ133505 (BjHD01), YS32 EF205404 (YS32), AF035832 (HB55), EF2053277 (HLD65), laboratory-acquired SEOV AF458104 (Ir461); SEOV strain U00465 (Houston), SEOV strain M34882 (Sapporo), SEOV strain U00463 (Egypt).
RESULTS
CHARACTERIZATION AND EVALUATION OF SEOV rNP EXPRESSED IN BACTERIA
We used pRSET B in the BL21 inducible system, in which the SEOV rNP of 430 amino acids expresses and purifies very well. The nucleotide sequence of the expression vector construct identified the expected sequence. SDS-PAGE revealed highly purified rNP with a molecular weight of about 51 kDa. The yield of SEOV rNP per 500 mL of E. coli cells ranged from 3 to 4 mg of protein.
The newly generated rNP was evaluated for use as a diagnostic antigen in an IgG ELISA. The reactivity patterns of rNP agreed with the reactivity patterns of Vero E6 cell-cultured authentic virus (data not shown). To analyze the cross-reactivity of ANDV virus with SEOV, rat sera were evaluated in an ELISA using both rNP antigens. The reactivity pattern of the ANDV rNP was different from the reactivity pattern of SEOV rNP (Figure 1). The OD values varied between different samples at the serum dilution (1:200) used and for the end-point titer values. Rat sera with low OD values could not be detected by ANDV rNP, whereas in sera with high IgG titers rNPs were not able to differentiate ANDV from SEOV infections serologically. Three SEOV RT-PCR positive rat samples with high OD values were indistinguishable for ANDV and SEOV rNPs.
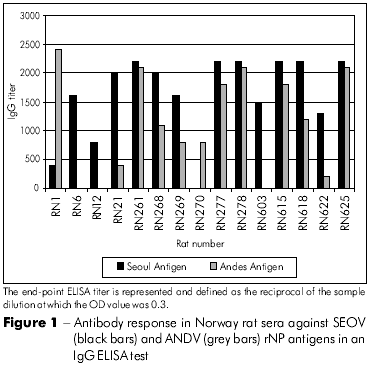
SCREENING OF RODENTS FOR THE PRESENCE OF VIRAL GENOME
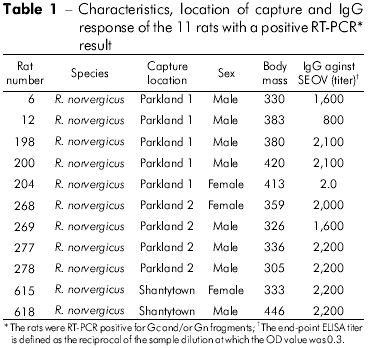
Of 151 rats captured in Buenos Aires City, 21 Norway rats (13.9%) were seropositive for SEOV rNP antigen. Serologically positive rats were tested for SEOV by RT-PCR using specific primers from two fragments of the M segment that encode Gn and Gc proteins. The viral genome was detected in the lungs of 11 of 21 seropositive rats (52.4%) captured in two parklands (Table 1). Eight out of 11 rats were males (72.7%), and all of them had adequate antibody responses. As expected, no product was amplified from the organs of the six seronegative rodents. In our ELISA tests, two of the Norway rats had high titers against the ANDV antigen and very low titers against the SEOV rNP antigen. We do not know if these rats had previous contact with infected Oligoryzomys spp rodents. Unfortunately, no viral genome was amplified from these animals.
GENETIC COMPARISON AND PHYLOGENETIC ANALYSIS
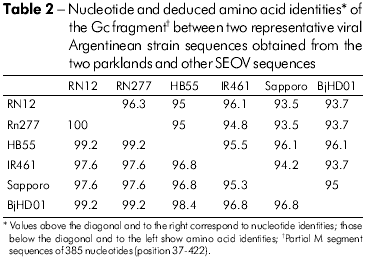
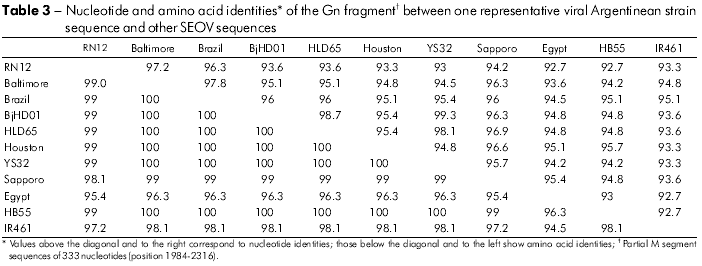
Among the 11 Gn amplified products, six cDNAs representative of two capture sites were sequenced. Pairwise comparison of sequences from Gn fragment (385 nt) among the three Norway rats (RN 12, RN 198 and RN 200) from one park and three from the second park (RN204, RN277, and RN278) showed 100% identity between the three rats from each group (Table 2). Representative rats from the two parks (RN12 and RN277) showed 96.3% nucleotide identity. Comparison of the deduced amino acid sequences of the six rats revealed 100% identity. Comparison of the Gn fragments from both rats with those of other representative SEOV showed the highest nucleotide identity with IR461 and HB55 Chinese strains (AF458104 and AF035832). Sequence comparisons of a 333-nt Gc fragment from one representative Argentinean rat (RN12) with other SEOV strains showed higher nucleotide identities with those strains from Baltimore, USA, and from Brazil (97% and 96%, respectively; Table 3). Phylogenetic analyses based on the 333-nt Gc fragment generated by the parsimonious method placed the SEOV RN 12 strain from Buenos Aires City together with the Baltimore and Brazil strains (Figure 2).
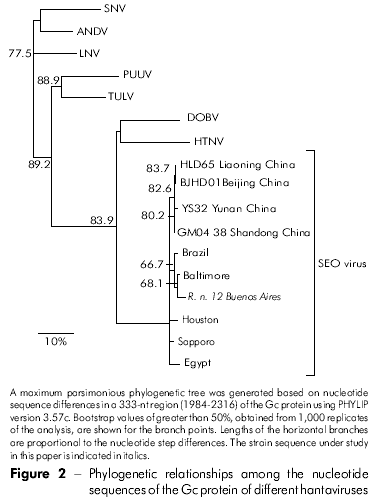
DISCUSSION
In this study, we report for the first time the detection of genetic material of SEOV in seropositive rats captured in Buenos Aires City, Argentina. The presence of SEOV in Buenos Aires City is not surprising, as previous studies have reported 31% seropositivity for SEOV23. Its reservoirs, R. norvegicus and R. rattus, are present in the country, and the distribution of this virus is reported worldwide. No seropositive black rats were detected; in addition, seropositive Norway rats were not equally distributed in the five different sites studied in Buenos Aires City, where the seroprevalence varied from 0% to 26.1%2. The serologic assay is an indirect measure of viral infection and is variably affected by the immune status of the host; however, it is the method most frequently used to evaluate hantavirus infection in the blood of rodents.
Earlier, we showed that ANDV rNP-based ELISA works well for the detection of virus-specific antibodies in patients and rodents with ANDV. In the present work, a serological assay was developed for use in the diagnosis of SEOV. We showed that the antigenicity of the complete SEOV rNP was strong enough to be utilized in an ELISA test. The ELISA based on the use of this antigen yielded results that were in good agreement with the results obtained by using cell culture fluids of SEOV. The assay could detect IgG antibody reactivity in rats and in control patient samples with SEOV infections from other parts of the world (data not shown). We also analyzed rat sera with low SEOV-ELISA values using ANDV rNP antigen, as previously described18, because sigmodontine-borne viruses were reported to be circulating around Buenos Aires City15. In a latitudinal study in natural areas of continental Chile, of 178 black rats studied, only one was found to be reactive in an ANDV-based ELISA serological test16.
The detection of antibodies does not necessarily indicate that the rodent is currently infected or infectious, but provides only indirect evidence of the current level of virus transmission. In areas where rats share habitat with sigmodontines, such as in the surroundings of Buenos Aires City, positive serologic results could represent the incidental spillover of AND-like virus. Thus, serum from rodents with antibodies to other non-pathogenic or pathogenic hantaviruses can test positive (i.e., cross-react) in the SEOV assay. For this reason, direct nucleic acid detection by RT-PCR has surfaced as the method-of-choice for a variety of viral pathogens. Successful viral genome amplifications were obtained from 11 seropositive rats. Our results confirm that SEOV is circulating in Buenos Aires City. This work constitutes direct evidence of current viral infection and indicates a high level of infected rats among seropositive Norway rat populations in the city because more than half of the seropositive rats carried detectable viral genomes. It is of interest that the newly described SEOV strain from Buenos Aires is most closely related to the wild type strains from Baltimore, USA, and from Brazil, both of which are associated with the Norway rat. The port of Buenos Aires is responsible for 80% of the transport of Argentine imports. Approximately 50% of the male Norway rats in Baltimore, Maryland, are infected with SEOV3,6. Studies with marked and released Norway rats in Baltimore demonstrate that male rats wounded between captures are more likely to become infected with SEOV upon recapture than are males that were not wounded between captures6. In our study, 72.7% of the rats with SEOV genome were males, although wounds were rarely observed in the captured rats. Among natural populations of Norway rats, males with more severe wounds are also more likely to shed SEOV and have viral RNA present in target tissues, such as the lungs, than are males with less severe or no wounds8.
SEOV has been reported to cause generally milder human disease than do HTNV and DOBV, but it is still responsible for 25% of the HFRS cases in Asia11. No human SEOV infections have been reported in Argentina. The presence of Norway rats with antibodies against SEOV is only one of many factors to consider when evaluating the risk of HFRS. Agents, reservoirs, the environment, and host factors interact to determine whether effective transmission, infection, and disease are likely. Agent factors include viral strain and the infecting dose of the particular hantavirus. Reservoir factors include active infection (vs. serologic evidence of past infection), the concentration of virus shed in the excreta, the density of rodents, and the frequency and spread of excretion. All of these factors should be considered when evaluating the significance of rodent serology data and when designing an appropriate preventive plan. Therefore, the significance of hantavirus RNA detection in rats as the causative agents for HFRS in Buenos Aires City remains unclear, and further serological surveys amongst healthy people or renal disease patients are needed, as well as environmental studies. Nevertheless, our results indicate that control strategies are needed to reduce the risk of rat-borne pathogen spillover to humans.
FINANCIAL SUPPORT
This work has been partially supported by the Government of Buenos Aires City.
REFERENCES
1 Billecocq A, Coudrier D, Boue F, Combes B, Zeller H, Artois M, et al. Expression of the nucleoprotein of the Puumala virus from the recombinant Semliki Forest virus replicon: characterization and use as a potential diagnostic tool. Clin Diagn Lab Immunol. 2003 Jul;10(4):658-63. [ Links ]
2 Cueto GR, Cavia R, Bellomo C, Padula PJ, Suarez OV. Prevalence of hantavirus infection in wild Rattus norvegicus and R. rattus populations of Buenos Aires City, Argentina. Trop Med Int Health. 2008 Jan;13(1):46-51. [ Links ]
3 Easterbrook JD, Kaplan JB, Glass GE, Pletnikov MV, Klein SL. Elevated testosterone and reduced 5-HIAA concentrations are associated with wounding and hantavirus infection in male Norway rats. Horm Behav. 2007 Nov;52(4):474-81. [ Links ]
4 Elgh F, Lundkvist A, Alexeyev OA, Stenlund H, Avsic-Zupanc T, Hjelle B, et al. Serological diagnosis of hantavirus infections by an enzyme-linked immunosorbent assay based on detection of immunoglobulin G and M responses to recombinant nucleocapsid proteins of five viral serotypes. J Clin Microbiol. 1997 May;35(5):1122-30. [ Links ]
5 Feldmann H, Sanchez A, Morzunov S, Spiropoulou CF, Rollin PE, Ksiazek TG, et al. Utilization of autopsy RNA for the synthesis of the nucleocapsid antigen of a newly recognized virus associated with hantavirus pulmonary syndrome. Virus Res. 1993 Dec;30(3):351-67. [ Links ]
6 Glass GE, Childs JE, Korch GW, LeDuc JW. Association of intraspecific wounding with hantaviral infection in wild rats (Rattus norvegicus). Epidemiol Infect. 1988 Oct;101(2):459-72. [ Links ]
7 Glass GE, Watson AJ, LeDuc JW, Childs JE. Domestic cases of hemorrhagic fever with renal syndrome in the United States. Nephron. 1994;68(1):48-51. [ Links ]
8 Hinson ER, Shone SM, Zink MC, Glass GE, Klein SL. Wounding: the primary mode of Seoul virus transmission among male Norway rats. Am J Trop Med Hyg. 2004 Mar;70(3):310-7. [ Links ]
9 Iversson LB, Rosa APA, Travassos da Rosa MDB, Lomar AV, Sasaki MGM, LeDuc JW. Human infection by hantavirus in southern and southeastern Brazil. Rev Assoc Med Bras. 1994 Apr-Jun;40(2):85-92. [ Links ]
10 Kallio-Kokko H, Vapalahti O, Hedman K, Brummer-Korvenkontio M, Vaheri A. Puumala virus antibody and immunoglobulin G avidity assays based on a recombinant nucleocapsid antigen. J Clin Microbiol. 1993 Mar;31(3):677-80. [ Links ]
11 Kim YS, Ahn C, Han JS, Kim S, Lee JS, Lee PW. Hemorrhagic fever with renal syndrome caused by the Seoul virus. Nephron. 1995;71(4):419-27. [ Links ]
12 LeDuc JW, Smith GA, Pinheiro FP, Vasconcelos PFC, Rosa EST, Maiztegui JI. Isolation of a Hantaan-related virus from Brazilian rats and serologic evidence of its widespread distribution in South America. Am J Trop Med Hyg. 1985 Jul;34(4):810-5. [ Links ]
13 Lee HW. Hemorrhagic fever with renal syndrome (HFRS). Scand J Infect Dis Suppl. 1982;36:82-5.
14 Lopez N, Padula P, Rossi C, Lazaro ME, Franze-Fernandez MT. Genetic identification of a new hantavirus causing severe pulmonary syndrome in Argentina. Virology. 1996 Jun;220(1):223-6. [ Links ]
15 Martinez VP, Colavecchia SB, Garcia Alay M, Suzuki B, Trincheri A, Busto S, et al. Hantavirus pulmonary syndrome in Buenos Aires Province. Medicina (B Aires). 2001;61(2):147-56. [ Links ]
16 Medina RA, Torres-Perez F, Galeno H, Navarrete M, Vial PA, Palma RE, et al. Ecology, genetic diversity, and phylogeographic structure of Andes virus in humans and rodents in Chile. J Virol. 2009 Mar;83(6):2446-59. [ Links ]
17 Nichol ST, Spiropoulou CF, Morzunov S, Rollin PE, Ksiazek TG, Feldmann H, et al. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science. 1993 Nov;262(5135): 914-7. [ Links ]
18 Padula P, Rossi CM, Della Valle MO, Martínez PV, Colavecchia SB, Edelstein A, et al. Development and evaluation of a solid-phase enzyme immunoassay based on Andes hantavirus recombinant nucleoprotein. J Med Microbiol. 2000 Feb;49(2):149-55. [ Links ]
19 Reynes JM, Soares JL, Hue T, Bouloy M, Sun S, Kruy SL, et al. Evidence of the presence of Seoul virus in Cambodia. Microbes Infect. 2003 Jul;5(9):769-73. [ Links ]
20 Schmaljohn CS, Hasty SE, Harrison SA, Dalrymple JM. Characterization of Hantaan virions, the prototype virus of hemorrhagic fever with renal syndrome. J Infect Dis. 1983 Dec;148(6):1005-12. [ Links ]
21 Schmaljohn CS, Hasty SE, Dalrymple JM, LeDuc JW, Lee HW, von Bonsdorff CH, et al. Antigenic and genetic properties of viruses linked to hemorrhagic fever with renal syndrome. Science. 1985 Mar;227(4690):1041-4.
22 Schmaljohn CS, Lee HW, Dalrymple JM. Detection of hantaviruses with RNA probes generated from recombinant DNA. Arch Virol. 1987;95(3-4):291-301. [ Links ]
23 Seijo A, Pini N, Levis S, Coto H, Deodato B, Cernigoi B, et al. Study of hantavirus seoul in a human and rodent population from a marginal area in Buenos Aires City. Medicina (B Aires). 2003;63(3):193-6. [ Links ]
24 Xiao SY, Leduc JW, Chu YK, Schmaljohn CS. Phylogenetic analyses of virus isolates in the genus Hantavirus, family Bunyaviridae. Virology. 1994 Jan;198(1):205-17.
25 Wang M, Rossi C, Schmaljohn CS. Expression of non-conserved regions of the S genome segments of three hantaviruses: evaluation of the expressed polypeptides for diagnosis of haemorrhagic fever with renal syndrome. J Gen Virol. 1993 Jun;74(Pt 6):1115-24. [ Links ]
26 Weissenbacher MC, Merani MS, Hodara VL, Villafane G, Gajdusek DC, Chu YK, et al. Hantavirus infection in laboratory and wild rodents in Argentina. Medicina (B Aires). 1990;50(1):43-6. [ Links ]
27 Zoller L, Yang S, Gott P, Bautz EK, Darai G. Use of recombinant nucleocapsid proteins of the Hantaan and nephropathia epidemica serotypes of hantaviruses as immunodiagnostic antigens. J Med Virol. 1993 Mar;39(3):200-7. [ Links ]
 Correspondência / Correspondence / Correspondencia:
Correspondência / Correspondence / Correspondencia:
Paula Julieta Padula Departamento de Virología,
Instituto Nacional de Enfermedades Infecciosas,
A.N.L.I.S. "Dr. Carlos G. Malbrán"
Av. Vélez Sarsfield 563 (1281)
Buenos Aires-Argentina
E-mail:ppadula@gmail.com
Phone/Fax: 5411 - 43013146
Recebido em / Received / Recibido en: 27/8/2009
Aceito em / Accepted / Aceito en: 30/3/2010
