










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.1 n.3 Belém dez. 2006
Análise estrutural da vegetação arbórea dos mangues no Furo Grande, Bragança, Pará
Structural analysis of mangrove tree vegetation in the channel Furo Grande in Bragança, Pará
Joaquim Augusto Souza de Seixas; Marcus Emanuel Barroncas Fernandes; Ederly Santos Silva
Universidade Federal do Pará. Campus de Bragança. Instituto de Estudos Costeiros. Laboratório de Ecologia de Manguezal. Bragança, Pará, Brasil (mebf@ufpa.br) (ederly.silva@bol.com.br)
RESUMO
O presente trabalho objetiva descrever os atributos estruturais dos bosques de mangue no Furo Grande, Bragança, Pará, enfatizando os padrões de distribuição espacial da vegetação, em 25 parcelas de 20x20 m. Para cada árvore foram retiradas as seguintes medidas: circunferência à altura do peito, altura total e largura da copa. Foram registrados 1.211 indivíduos de duas espécies, Rhizophora mangle e Avicennia germinans,distribuídos em três sítios. No Sítio no1, dominado por A. germinans,foram medidas 255 árvores de A. germinans (93,07%) e 19 de R. mangle (6,93%), totalizando 274 indivíduos. O Sítio no2, dominado por R. mangle, apresentou 382 árvores de R. mangle (90,31%) e 41 de A. germinans (9,69%), totalizando 423 indivíduos. Já o Sítio no3 apresentou 269 árvores de A. germinans (52,33%) e 245 de R. mangle (47,67%), totalizando 514 indivíduos. A espécie mais abundante neste estudo foi R. mangle com 646 indivíduos. A análise dos parâmetros estruturais apresentou diferença significativa entre DAP x Densidade, DAP x Altura e Largura da Copa x DAP Considerando os três sítios, os valores de Densidade apontaram o Sítio no1 (dominado por A. germinans) como o de maior porte. Essa diferença é corroborada pelos maiores valores médios de altura e DAP das árvores nesse sítio. A diferença nas características estruturais e composição florística desses bosques na área de estudo parece indicar que esses fatores bióticos podem estar sendo influenciados por outros fatores bióticos, como competição e sucessão natural dos indivíduos senescentes.
Palavras-chave: Atributos estruturais. Manguezal. Furo Grande. Bragança. Pará.
ABSTRACT
The present study aims to describe the structural attributes of the mangrove stands at the Furo Grande, Bragança, Pará, emphasizing the patterns of spatial distribution of the vegetation in 25 quadrats of 20x20 m. For every tree inside the plots the following measures were taken: Diameter at Breast Height (DBH), Height and Crown Width. Measurements were taken from 1211 individuals of two species, Rhizophora mangle and Avicennia germinans, distributed between three sites. At Site no1, dominated by A. germinans, 255 trees of A. germinans (93.07%) were measured and 19 of R. mangle (6.93%), totalling 274 individuals. Site no2, dominated by R. mangle, presented 382 trees of R. mangle (90.31%) and 41 of A. germinans (9.69%), totalling 423 individuals. Site no3 presented 269 trees of A. germinans (52.33%) and 245 of R. mangle (47.67%), totalling 514 individuals. The most abundant species in the present study was R. mangle with 646 individuals. The analysis of the structural parameters presented significant differences between DBH x Density (F = 6.07; gl=24; p<0.05), DBH x Height (F=18.70; gl=24; p<0.001) and Crown Width x DBH (F=47.15; gl=23; p<0.001). Of the three sites, density values showed Site no1 (dominated by A. germinans)as the most developed. This difference is corroborated by high mean values of Height and DBH. The results on structural attributes and floristic composition of mangrove stands at the Furo Grande seem to indicate that these biotic factors can be influenced by other biotic factors, such as competition and natural succession of senescent individuals.
Keywords: Structural attributes. Mangal. Bragança Peninsular. Bragança. Pará.
INTRODUÇÃO
No Brasil, os manguezais cobrem cerca de 20.000 km2 (YOKOYA, 1995), o que equivale a mais de 12% do total de áreas cobertas por este ecossistema em todo o planeta, correspondendo às maiores áreas de manguezal encontradas na costa norte, que vai do Oiapoque, Amapá, até o Golfo maranhense, Maranhão (FERNANDES, 2003).
O estuário do rio Caeté situa-se dentro da segunda maior região de manguezal do norte do Brasil, com uma abrangência de aproximadamente 220 km2 (WOLFF; KOCH; ISSAC, 2000). Esta região, localizada a 200 km a noroeste de Belém, caracteriza-se pela ação de macromarés que podem variar de 2,5 a 5,5 m no médio estuário (KOCH, 1999). Apresenta, também, duas estações climáticas bem definidas, sendo uma estação seca, que se prolonga de julho a dezembro, e uma chuvosa, de janeiro a junho. Além da pluviosidade, existem outros fatores abióticos que influenciam no sucesso do estabelecimento dos bosques de mangue.
Para Alongi (1989), a existência de manguezais depende principalmente da existência de temperatura atmosférica superior a 15oC e a presença de águas rasas. Este autor afirma, também, que para uma floresta de mangue se estabelecer em determinadas áreas é necessário que ela esteja protegida contra a ação das ondas, que poderiam impedir a fixação e desenvolvimento adequado das plântulas, considerando a mobilidade do substrato que o manguezal costuma colonizar.
A variabilidade estrutural das florestas de mangue na costa brasileira foi descrita em termos de unidades fisiográficas por Schaeffer-Novelli et al. (1990). De acordo com estes autores, a pluma do Amazonas carreia grande quantidade de sedimento, influenciando os padrões estruturais dos manguezais na região amazônica, principalmente no estado do Amapá. De acordo com Tomlinson (1986), os manguezais dividem uma interessante mistura de atributos. Essas diferentes características são, geralmente, consideradas como respostas aos fatores ambientais que influenciam seu desenvolvimento. A arquitetura da floresta, por exemplo, é influenciada pela magnitude e periodicidade desses fatores abióticos, os quais variam nas mais diferentes escalas.
Na costa norte brasileira são poucos os estudos que enfocam a estrutura e dinâmica das maiores e mais desenvolvidas florestas de mangue do país (DAMÁSIO, 1980a, 1980b; SCHAEFFER-NOVELLI; CINTRÓN, 1988; ALMEIDA, 1996; PROST; RABELO, 1996; BASTOS; LOBATO, 1996; FERNANDES, 1997; MENEZES; BERGER; WORBES, 2003). Em sendo assim, este trabalho tem o intuito de descrever os atributos estruturais de parte da floresta de mangue no Furo Grande, Bragança, Pará, dando ênfase aos padrões de distribuição espacial da vegetação local.
MATERIAL E MÉTODOS
Área de estudo
A planície costeira bragantina, nordeste do Pará, abrange a faixa costeira do município de Bragança, região que se estende da ponta do Maiaú até a foz do rio Caeté, perfazendo uma área de aproximadamente 1.570 km2 (SOUZA FILHO, 1995). Esta região possui uma linha costeira de aproximadamente 40 km, caracterizada pela presença de rios, manguezais e planaltos rebaixados. O presente trabalho foi realizado no Furo Grande, na península de Ajuruteua, próximo à cidade de Bragança, entre as coordenadas 00o50'19,5"S e 46o38'14,9"W (Figura 1). O manguezal estudado localiza-se ao lado esquerdo da rodovia PA-458, que liga Bragança à vila de Ajuruteua.
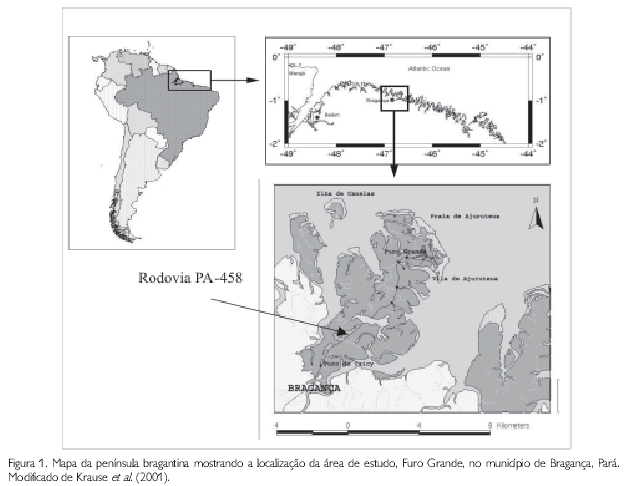
Fatores abióticos
O clima da região bragantina, assim como toda a costa nordeste do Pará, pode ser classificado como tropical quente e úmido (CRITCHFIELD, 1968). A pluviosidade média anual desta região é de 2.000 mm, com umidade relativa do ar oscilando entre 80 e 91% (MARTORANO et al.,1993).
Ao longo de dezessete anos de monitoramento, a região apresenta uma temperatura média de 25,9oC, com período de estiagem de agosto a dezembro. O regime de marés é semidiurno, ocorrendo subida e descida duas vezes ao dia durante um período de aproximadamente 6,2 h, além de apresentar um sistema de macromarés (4 a 5 m) com o ciclo total de 24,5 h. A descarga do rio Caeté é de aproximadamente 180 m3/s durante a estação da chuva e 0,3 m3/s durante o período de seca (SCHWENDENMANN, 1998).
A floresta de mangue na península bragantina é inundada inteiramente apenas nas marés de sizígia, enquanto os canais e furos dos manguezais são alagados diariamente. A energia das marés altas e as fortes correntes, durante as marés de sizígia, carreiam grande quantidade de sedimento, especialmente nas partes externas dos estuários (WOLFF; KOCH; ISAAC, 2000). No estuário e nos canais com maiores marés, a velocidade das correntes alcança valores máximos de aproximadamente 1,5 m.s-1 durante as marés de sizígia (COHEN, 1998).
Segundo Figueira (2002), a temperatura da área de estudo não varia muito, permanecendo entre 25 e 30oC ao longo do ano, o mesmo ocorrendo para os valores de pH, que oscilam entre 6 e 7. A salinidade apresenta a maior flutuação, atingindo valores entre 5 e 35.
Vegetação característica
As espécies características de mangue que ocorrem na área de estudo do Furo Grande são Rhizophora mangle (mangue vermelho) e Avicennia germinans (mangue preto ou siriúba), que ocupam a maior parte da área de estudo. Há ocorrência de Laguncularia racemosa (mangue branco ou tinteira), observada apenas na borda da floresta, além de um estrato herbáceo composto basicamente de Sesuvium portulacastrum.
Procedimento
Os métodos descritos abaixo foram baseados em Schaeffer-Novelli e Cintrón (1986) e usados para avaliar as características estruturais da floresta de mangue no Furo Grande. Vinte e cinco parcelas de 20x20 m, totalizando um hectare (10.000 m2) foram distribuídas na área de estudo. A vegetação é separada por pequenos canais internos, formando, assim, três paisagens bastante diferenciadas. As parcelas foram distribuídas de forma que as três paisagens fossem descritas quanto aos seus atributos estruturais. No bosque mais próximo à margem do Furo Grande, aqui denominado Sítio no3, foram medidas 9 parcelas. Nos bosques mais internos, aqui denominados Sítio no2 e Sítio no1, foram medidas 8 parcelas em cada sítio. Em cada parcela foram mensuradas todas as árvores com o mínimo de 10 cm de circunferência à altura do peito (CAP). Para cada árvore foram retiradas as da circunferência à altura do peito, altura total e largura da copa. A altura foi estimada com o uso de um telêmetro (30 m de alcance), somando-se a altura do observador em linha reta com o ponto visível mais alto da árvore, enquanto que a largura da copa foi medida esticando-se uma trena (50 m) entre as porções laterais mais externas da copa.
Análise de dados
Os dados obtidos basearam a estimativa dos seguintes parâmetros estruturais, com fundamento no estudo de Schaeffer-Novelli e Cintrón (1986): diâmetro à altura do peito (DAP), freqüência (F = presença/ausência da espécie por parcela), densidade (De = número de indivíduos por parcela), dominância (Do = área basal de cada espécie por parcela), valor de cobertura (VC = De + Do) e valor de importância (VI = F + De + Do).
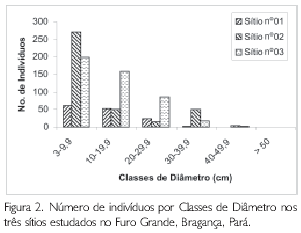
Os indivíduos registrados nos três sítios de trabalho foram subdivididos em seis classes de diâmetro, com o objetivo de analisar o desenvolvimento dos bosques estudados. As classes utilizadas foram: a) 3 a 9,9 cm; b) 10 a 19,9 cm; c) 20 a 29,9 cm; d) 30 a 39,9 cm; e) 40 a 49,9 cm e f) maior que 50 cm.
Os valores encontrados a partir da estimativa dos parâmetros estruturais dos três sítios de trabalho foram comparados entre si no intuito de avaliar se a diferença desses valores apresentava um nível de significância mínimo, no intervalo de 5%, através da Análise de Variância (ANOVA - um fator). A regressão linear foi utilizada para analisar a arquitetura dos bosques através da relação dos valores de DAP altura total (AT), largura da copa (LC) e De. Todas as análises estatísticas foram desenvolvidas no pacote BioEstat 3.0 (AYRES, M.; AYRES JUNIOR; SANTOS, 2003).
RESULTADOS
Foram registrados 1.211 indivíduos, pertencentes a duas espécies, Rhizophora mangle e Avicennia germinans,distribuídos nos três sítios. No Sítio no1, constituído basicamente de A. germinans, foram mensuradas 255 árvores de A. germinans (93,07%) e 19 de R. mangle (6,93%), totalizando 274 indivíduos. O Sítio no2, composto principalmente por R. mangle,apresentou um total de 382 árvores de R. mangle (90,31%) e 41 árvores de A. germinans (9,69%), perfazendo um total de 423 indivíduos. Já o Sítio no3 apresentou 269 árvores de A. germinans (52,33%) e 245 de R. mangle (47,67%), gerando um total de 514 indivíduos.
Em relação à quantidade de indivíduos mortos em cada sítio de trabalho, no Sítio no1 foram registrados 28 indivíduos de A. germinans e 3 indivíduos de R. mangle,enquanto no Sítio no2 foram registrados 5 de A. germinans e 22 de R. mangle. O Sítio no3 foi o que apresentou o maior número de árvores mortas, sendo 67 de A. germinans e 8 de R. mangle.
A Figura 2 estabelece a relação entre o número de indivíduos e as classes de diâmetro, mostrando que o Sítio no2 apresenta o maior número de indivíduos (n = 272) pertencentes à menor classe. Referente à segunda classe, o maior número de indivíduos foi encontrado no Sítio no3 (n=158), enquanto os outros sítios apresentaram a mesma quantidade de indivíduos (n = 54). As duas últimas classes não são tão expressivas quanto ao número de indivíduos, ou seja, da quinta classe, o Sítio no2 apresentou cinco indivíduos e o Sito no3 apenas três. Da última classe, indivíduos que possuem DAP maior do que 50 cm, foi registrada somente uma árvore no Sítio no2.
A freqüência relativa (FR) das espécies botânicas em todos os sítios de trabalho apresentou valores semelhantes, ou seja, R. mangle e A. germinans foram registradas pelo menos uma vez em cada parcela. O Sítio no1, por sua vez, mostrou uma pequena variação, registrando 46,67% para Rhizophora, isto é, em algumas parcelas não houve registro de indivíduos desse tipo de mangue (Tabela 1).
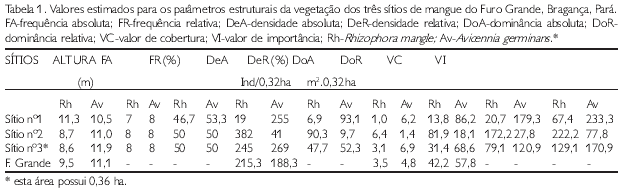
A espécie com maior densidade foi R. mangle (215,3 ind.ha), enquanto que A. germinans apresentou o maior valor de dominância (4,8 m2.ha) para o Furo Grande. O Sítio no3 apresentou os maiores valores para densidade e dominância (Tabela 1).
O valor de cobertura (VC) e o valor de importância (VI) também apresentaram o mesmo padrão que os parâmetros anteriores para os três sítios, ou seja, os maiores valores para o Sítio no1 e o Sítio no3 são de A. germinans.VC-179,3 e VI-233,3 e VC-120,9 e VI-170,9, respectivamente (Tabela 1).
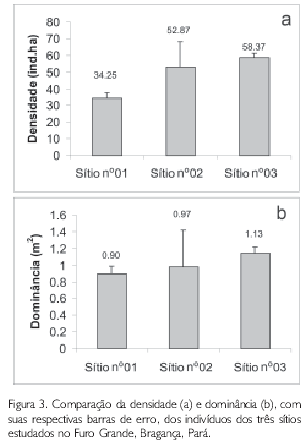
Através da análise de variância, foi testada a diferença entre os valores obtidos para a densidade e dominância nos três sítios de trabalho. O resultado desta análise mostrou que não há diferença significativa (ANOVA, p>0,05) entre os sítios estudados. A Figura 3 demonstra que tanto para densidade quanto para a dominância, os maiores valores são encontrados no Sítio no3.
A Figura 4 mostra as relações significativas entre os atributos estruturais das duas espécies presentes nas parcelas estudadas nos três sítios do Furo Grande, analisadas através da regressão linear. A relação entre o DAP e a densidade absoluta para os indivíduos de A. germinans foi significativa (F=6,07; gl=24; p<0,05), ao passo que a relação entre diâmetro e densidade absoluta para R. mangle não foi significativa (p>0,05). O mesmo resultado foi obtido para a relação entre o DAP e a altura foi analisada, pois somente para A. germinans a correlação foi significativa (F = 18,70; gl = 24; p<0,001). Já a relação entre a altura e a densidade absoluta não foi significativa (p>0,05) para ambas as espécies. Por fim, a relação largura da copa e DAP foi significativa somente para R. mangle (F=47,15; gl=23; p<0,001).
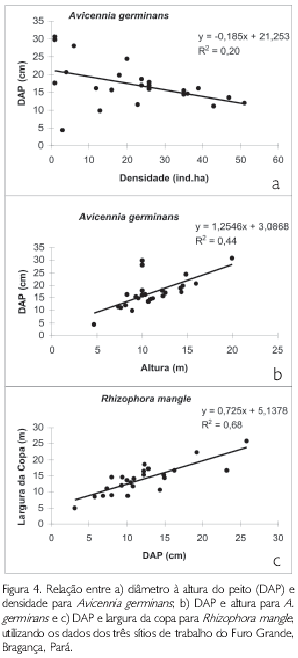
DISCUSSÃO
De acordo com os trabalhos realizados sobre a estrutura dos bosques de mangue na região amazônica, a paisagem formada pelo manguezal é principalmente dominada pelos gêneros Rhizophora, Avicennia e Laguncularia (ALMEIDA, 1996; PROST; RABELO, 1996; BASTOS; LOBATO, 1996; FERNANDES, 1997, 2002; MENEZES; BERGER; WORBS, 2003). Os bosques estudados no Furo Grande, embora apresentem Laguncularianas bordas, possuem Avicennia e Rhizophora como os principais formadores da paisagem local, com o Sítio no1 e Sítio no 2 dominados por estes gêneros, respectivamente, e o Sítio no3 possuindo uma formação mista desses dois gêneros.
Segundo Schaeffer-Novelli e Cintrón (1986), os bosques passam, durante seu desenvolvimento, por uma fase em que a área está ocupada por uma grande densidade de árvores de diâmetros reduzidos e, posteriormente, a uma fase de maior amadurecimento, quando o domínio é feito por poucas árvores de grande porte. Nos bosques estudados, mais de 50% dos indivíduos possuem DAP bastante reduzido, pertencendo à primeira classe de diâmetro aqui estabelecida (3 a 9,9 cm), mostrando que os bosques possuem uma alta densidade de indivíduos de diâmetro reduzido. No entanto, considerando a presença de árvores adultas, com porte de até 25 m de altura e DAP em torno de 45 cm, estes registros podem indicar a presença de um bosque adulto, mas em processo de substituição de indivíduos senescentes por indivíduos jovens. A presença de muitos indivíduos mortos nas parcelas estudadas parece corroborar a idéia de que o bosque está em processo, já adiantado, de substituição desses indivíduos.
Considerando os estudos que utilizaram o mesmo método de parcela para a costa norte, os valores de densidade encontrados para o Furo Grande são equivalentes aos estimados por Bastos e Lobato (1996) para a praia do Crispim e Algodoal, no nordeste do Pará, cujos valores atingem 595,2 e 364,8 ind.0,32ha, respectivamente. Já os dados produzidos por Menezes, Berger e Worbs (2003) na península bragantina, nas localidades Acarajó, Furo do Chato e Furo Grande, são inferiores aos desse estudo (195,2; 118,4 e 201,6 ind.0,32ha, respectivamente), enquanto a dominância apresenta valores similares, principalmente no Furo do Chato (9,7 m2.0,32ha). Contudo, ainda são poucos os dados publicados para efeito comparativo no litoral Amazônico, mais raros ainda se realizados utilizando metodologia diferente.
Segundo Almeida (1996), os manguezais localizados mais próximos ao estuário são mais densos e ricos em espécies do que aqueles sob influência marinha, em função do estresse causado pela alta salinidade. A área do Furo Grande apresenta salinidade variável ao longo do ano (de 5 a 35) (FIGUEIRA, 2002), sugerindo que os sítios enfocados no presente estudo estão sob maior influência da salinidade somente durante o período seco, quando os níveis de sal alcançam os maiores valores. Sendo assim é razoável pensar que os valores de densidade encontrados não parecem estar sendo influenciados pela salinidade, mas muito provavelmente por outro fator, como ocupação de antigas clareiras provocadas pela queda de árvores senescentes.
Segundo Schaeffer-Novelli e Cintrón (1986), a densidade de um bosque de mangue é uma função de sua idade e desenvolvimento. Se forem considerados os três bosques no Furo Grande, o Sítio no3 (dominado por A. germinans) é o mais desenvolvido por apresentar o menor valor de densidade e o maior de dominância, além de apresentar os maiores valores de altura média e DAP.
Em considerando o valor de cobertura e o valor de importância registrado para os três sítios, o Sítio no1 (A. germinans) e o Sítio no2 (R. mangle) apresentaram valores maiores do que 90%. Isto implica na presença da quase totalidade de indivíduos das respectivas espécies dominantes, ou seja, são quase bosques puros, sendo a presença de outras espécies bastante esparsa. Já o Sítio no3 possui uma característica bastante diferenciada, apresentando percentuais equivalentes para as duas espécies de mangue. Este fato atesta a grande discrepância nas características estruturais e composição desses bosques na área de estudo do Furo Grande, o que neste caso parece receber pouca influência dos fatores abióticos, principalmente salinidade.
No estudo desenvolvido por Menezes, Berger e Worbs (2003) na península bragantina, também foi sugerida uma baixa influência dos fatores abióticos sobre a estrutura dos bosques. De fato, estes autores sugerem que os bosques investigados apresentam diferentes estágios de desenvolvimento e as diferenças em densidade e dominância apresentadas poderiam ser fruto de um processo de declínio natural na densidade populacional ao invés do efeito direto de fatores abióticos. Contudo, segundo Tomlinson (1986), os mangues compartilham uma mistura interessante de atributos. Eles apresentam características pronunciadas de espécie pioneiras, maximizando o aumento da população e mantendo uma dada área com o máximo da sua capacidade de suporte. Essas características são consideradas uma resposta aos fatores ambientais que influenciam o desenvolvimento do bosque desde os estágios iniciais até à maturidade.
Por fim, segundo Fernandes (2003), a arquitetura da floresta, por exemplo, é influenciada pela magnitude e periodicidade de fatores ambientais que variam de região para região ou até dentro de uma mesma área. Assim, os resultados obtidos no presente trabalho parecem indicar que esses fatores bióticos podem estar sendo influenciados por outros fatores bióticos, como competição e sucessão natural dos indivíduos senescentes. Por outro lado, a distribuição das diferentes partes que formam o mosaico do solo pode também ter afetado tal distribuição vegetacional, com a formação de distintos bosques de mangue, haja vista já existir fortes evidências mostrando diferenças significativas entre solos de mangue caracterizados pela presença de diferentes espécies de mangue (LACERDA; ITTEKKOT; PATCHINEELAM, 1995).
CONCLUSÕES
Somente duas espécies de mangue foram registradas nas parcelas dos três sítios de trabalho: R. mangle e A. germinans, muito embora haja ocorrência de L. racemosa nas bordas da área de estudo, no Furo Grande. Os indivíduos de A. germinans possuem estrutura mais desenvolvida do que os de R. mangle.
O Sítio no1 é o bosque mais maduro, com árvores mais desenvolvidas e com menor valor de densidade. O Sítio no2 é o bosque de menor porte e com maior densidade de indivíduos jovens.
A seqüência estudada apresenta três paisagens distintas de mangue, sendo uma dominada por Avicennia germinans (Sítio no1), outra dominada por Rhizophora mangle (Sítio no2) e uma terceira faixa considerada mista, por ser dominada por ambas as espécies (Sítio no3).
REFERÊNCIAS
ALMEIDA, S. S. 1996. Estrutura e florística em áreas de manguezais paraenses: evidências da influência do estuário amazônico. Bol. Mus. Para. Emilio Goeldi, v. 8, p. 93-100.
ALONGI, D. M. 1989. The role of soft-bottom benthic communities in Tropical Mangrove and Coral Reef Ecosystems. Critical Reviews in Aquatic Sciences, v. 1, n. 2, p. 243-280.
AYRES, M.; AYRES JUNIOR, M.; SANTOS, A. S. 2003. BioEstat 3.0: aplicações estatísticas nas áreas das ciências biológicas e médicas. Belém: Sociedade Civil Mamirauá. 290 p.
BASTOS, M. N. C.; LOBATO, L. C. B. 1996. Estudos fitossociológicos em áreas de bosque de mangue na praia do Crispim e Ilha de Algodoal-Pará. Bol. Mus. Para. Emilio Goeldi, v. 8, p. 1 57-1 67.
COHEN, M. C. L. et al. 1998. Factors influencing the variability of Mg, Ca and K in waters of a mangrove creek, in Bragança, North Brazil. Mangroves and Salt Marshes, v. 44, p. 1-7.
CRITCHFIELD, H. J. 1968. General Climatology. New Delhi: Prentice Hall. 420 p.
DAMÁSIO, E. 1980a. Contribuição ao conhecimento da vegetação dos manguezais da Ilha de São Luis. Bol. Lab. Hidrob., v. 3, p. 1756. Part I.
DAMÁSIO, E. 1980b. Contribuição ao conhecimento da vegetação dos manguezais da Ilha de São Luis. Bol. Lab. Hidrob., v. 3, p. 5776. Part II.
FERNANDES, M. E. B. 1997. The ecology and productivity of mangroves in the Amazon region, Brazil. 214 f. Tese (Doutorado) - University of York, England.
FERNANDES, M. E. B. 2002. Structural analysis of Rhizophora, Avicennia, and Laguncularia forests on Maracá Island, Amapá, Brazil. In: LIEBEREI, R. et al. (Ed.). Neotropical Ecosystems: proceedings of the German-Brazilian Workshop. p.565-572. Hamburg 2000. GKSS-Geesthacht. 1 CD-ROM.
FERNANDES, M. E. B. 2003. Produção Primária: serapilheira. In: FERNANDES, M. E. B. (Org.). Os manguezais da costa norte brasileira. São Luís-MA: Fundação Rio Bacanga. p. 61-78.
FIGUEIRA, E. A. G. 2002. Caracterização da comunidade macrobentônica dos manguezais do Furo Grande, Bragança, Pará. 109 f. Dissertação (Mestrado) - Universidade Federal do Pará.
KOCH, V. 1999. Epibenthic production and energy flow in the Caeté mangrove estuary, North Brazil. 96 f. Tese (Doutorado) - Universidade de Bremen, Alemanha.
KRAUSE, G. et al. 2001. Spatial Patterns of Mangrove Ecosystems: the bragantinian mangroves of Norther Brazil. Ecotropica, v. 7, p. 93-170.
LACERDA, L. D.; ITTEKKOT, V.; PATCHINEELAM, S. R. 1995. Biogeochemistry of mangrove soil organic matter: a comparison between Rhizophora and Avicennia soils in South-eastern Brazil. Coastal and Shelf Science, Estuarine, v. 40, p. 713-720.
MARTORANO, L. G. et al. 1993. Estudos climáticos do Estado do Pará, classificação climática (Köppen) e deficiência hídrica (Thornthwhite, Mather). Belém: SUDAM/EMBRAPA, SNLCS. 53 p.
MENEZES, M. P. M.; BERGER, U.; WORBES, M. 2003. Annual growth rings and long-term growth patterns of mangrove trees from the Braganca's peninsula, north Brazil. Wetlands Ecology and Management, v. 11, p. 233-242.
PROST, M. T. R. C.; RABELO, B. V. 1996. Variabilidade fito-espacial de manguezais litorâneos e dinâmica costeira: exemplos da Guiana Francesa, Amapá e Pará. Bol. Mus. Para. Emilio Goeldi, v. 8, p. 101-121.
SCHAEFFER-NOVELLI, Y.; CINTRÓN, G. M. 1986. Guia para estudo de áreas de manguezal: estrutura, função e flora. [S.l.]: Caribbean Ecological Research. 186 p.
SCHAEFFER-NOVELLI, Y.; CINTRÓN, G. M. 1988. Expedição Nacional aos Manguezais do Amapá, Ilha de Maracá. [S.l.:s.n.]. 99 p. Relatório Técnico- Cons. Nac. Desenv. Científico e Tecnológico- CNPq. Coordenação de Ciências Biológicas.
SCHAEFFER-NOVELLI, Y. et al. 1990. Variability of mangrove ecosystems along the Brazilian coast. Estuaries, v. 13, n. 2, p. 204-218.
SCHWENDENMANN, L. 1998. Tidal and seasonal variations of soil and water properties in a Brazilian mangrove ecosystem. Dissertação (Mestrado) - University of Karlsruhe, Karlsruhe.
SOUZA FILHO, P. W. M. 1995. A planície costeira bragantina (NE do PA): influência das variações do nível do mar na morfoestratigrafia costeira durante o Holoceno. 123 f. Dissertação (Mestrado) - Universidade Federal do Pará.
TOMLINSON, P. B. 1986. The botany of mangroves. Cambridge: Cambridge University Press. 419 p.
WOLFF, M.; KOCH, V.; ISAAC, V. 2000. A trophic flow model of the Caeté Mangrove Estuary (North Brazil) with considerations for the sustainable use of its resources. Estuarine Coastal. Shelf Science. v. 50, p. 789-803.
YOKOYA, N. S. 1995. Distribuição e Origem. In: SCHAEFFER-NOVELLI, Y. (Org.). Manguezal Ecossistema entre a terra e o mar. São Paulo: Caribbean Ecological Research. p. 9-12.
 Endereço para correspondência:
Endereço para correspondência:
Editora do Museu Paraense Emílio Goeldi
Av. Magalhães Barata, 376
São Braz – CEP 66040-170
Caixa Postal 399
Telefone/fax: 55-91-3219-3317
E-mail: boletim@museu-goeldi.br
Recebido: 10/03/2005
Aprovado: 27/03/2006
