










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.3 n.3 Belém dez. 2008
The skull and abdominal skeleton of Stenocercus dumerilii (Steindachner, 1867) (Reptilia: Squamata: Iguania)
Crânio e esqueleto abdominal de Stenocercus dumerilii (Steindachner, 1867) (Reptilia: Squamata: Iguania)
Emil José Hernández-RuzI; Teresa Cristina Sauer Avila-PiresII
IUniversidade Federal do Pará. Instituto de Ciências Biológicas. Laboratório de polimorfismo de DNA. Belém, Pará, Brasil (emilhjh@yahoo.com)
IIMuseu Paraense Emílio Goeldi. Coordenação de Zoologia. Belém, Pará, Brasil (avilapires@museu-goeldi.br)
ABSTRACT
The objective of this study is to describe the skull and abdominal skeleton of the lizard Stenocercus dumerilii (Steindachner, 1867), with emphasis on those characters used in phylogenetic studies involving the genus Stenocercus. The peculiar shape of the head (pyramidal, with a distinct canthal-superciliary ridge that ends in a prominent postsupraciliary scale) results in part from the shape of the prefrontal and postfrontal bones, both of which bear protuberances projecting towards the orbital cavity. The postfrontal bone represents the highest point of the cranium. Contrary to what has been observed in other species of Stenocercus and in most Tropiduridae, the studied specimens ofS. dumerilii presented a single pair of xiphisternal inscriptional ribs (followed by a pair of posterior xiphisternal processes). Besides, they presented five pairs of postxiphisternal inscriptional ribs widely separated ventrally. One specimen (of two studied) showed the pineal foramen completely enclosed by the frontal instead of in frontoparietal suture.
Keywords: Iguania. Cranial osteology. Abdominal skeleton. Stenocercus dumerilii.
RESUMO
O presente estudo tem por objetivo descrever o crânio e o esqueleto abdominal do lagarto Stenocercus dumerilii (Steindachner, 1867), com ênfase nos caracteres utilizados em estudos filogenéticos incluindo o gênero Stenocercus. A forma peculiar da cabeça (piramidal, com uma crista supraciliar bem demarcada e terminando em uma escama pós-supraciliar proeminente) resulta em parte do formato dos ossos pré-frontal e pós-frontal, ambos com protuberâncias que se projetam em direção à cavidade orbital. O osso pós-frontal representa o ponto mais alto da cabeça. Diferentemente do que foi encontrado até o momento nas espécies de Stenocercus e na maioria dos Tropiduridae, os exemplares estudados de S. dumerilii apresentaram um único par de costelas xifisternais (seguido por um par de processos posteriores do xifisterno). Ademais, apresentaram cinco pares de costelas pós-xifisternais amplamente separadas na região ventral. De dois exemplares estudados, em um deles o forâmen pineal é totalmente envolvido pelo osso frontal, ao invés de na sutura entre o frontal e o parietal.
Palavras-chave: Iguania. Osteologia craniana. Esqueleto abdominal. Stenocercus dumerilii.
INTRODUCTION
Stenocercus dumerilii (Figure 1) is a lizard with cryptic coloration and behavior, which inhabits eastern Pará (Cunha, 1981; Avila-Pires, 1995) and western Maranhão states (Andrade et al., 2003; Nogueira & Rodrigues, 2006), in Brazil. Its distribution is widely coincident with the Belém area of endemism, a biogeographic unit mainly defined on the basis of butterflies, birds and primates (see Silva et al., 2005 for a brief characterization of Amazonian areas of endemism). This is by far the most threatened area in Amazonia, with 65% of deforested area in numbers of early 2005 (Silva et al., 2005). Although Stenocercus dumerilii occurs in areas of secondary forest (Cunha, 1981), even such areas are now disappearing, which caused the species to be considered "endangered" in the list of endangered species of the state of Pará (SEMA-PA, 2007).

Stenocercus Duméril & Bibron, 1837 is part of the Tropiduridae sensu Frost et al. (2001) or the Iguanidae, Tropidurinae*, Tropidurini sensu Schulte et al. (2003). According to both papers it is the sister taxon of the Tropidurus group (sensu Frost, 1992). It is a complex genus, with 61 species recognized at present (Torres-Carvajal, 2007a, b), most of which occur in the Andean region. Stenocercus dumerilii was described by Steindachner (1867) in the genus Ophryoessoides Duméril, 1851, which until then included only the type-species, O. tricristatus Duméril, 1851. Boulenger (1885) synonymised Ophryoessoides and five other genera with Leiocephalus Gray, 1827. Etheridge (1966) restricted the name Leiocephalus to the West Indian species and allocated the remaining species in Ophryoessoides, the oldest available generic name. Since his study focused on the West Indian group, no detailed analysis of the 'Ophryoessoides' species was done. Fritts (1974) redefined Ophryoessoides and Stenocercus, allocating some species of Ophryoessoides to Stenocercus. Frost (1992), however, demonstrated that some species of Ophryoessoides sensu Fritts (1974) were more closely related to some Stenocercus species. He therefore postulated the synonymization of Ophryoessoides, as well as of Proctotretus Duméril & Bibron, 1837, with Stenocercus. Cadle (1991) and Avila-Pires (1995) followed Frost's (1992) proposal, emphasizing the need of more studies on the group. Some species within Stenocercus, however, distinguished by enlarged cephalic scales, enlarged supraoculars and keeled ventrals, were still informally recognized as the 'Ophryoessoides group' (Cadle, 1998, 2001). Recently, Torres-Carvajal et al. (2006) performed a phylogenetic analysis of Stenocercus using mtDNA sequence data, and Torres-Carvajal (2007a) presented a combined morphological-molecular phylogenetic analysis. This latter analysis included data on external morphology from all but two of the 61 known species, osteology from 46 species, and mtDNA from 35 species. Both studies recovered two main clades composing the genus, one of which nested the species previously in Proctotretus and Ophryoessoides (sensu Fritts, 1974), amid others. Stenocercus dumerilii (external morphology and osteology studied) was recovered by Torres-Carvajal (2007a) as the sister taxon of S. tricristatus (external morphology) and these two as sisters of the clade S. sinesaccus Torres-Carvajal, 2005 (S. caducus (Cope, 1862) - S. prionotus Cadle, 2001). The two species not included in the study, S. quinarius Nogueira & Rodrigues, 2006 and S. squarrosus Nogueira & Rodrigues, 2006, most similar in external morphology to S. tricristatus and S. dumerilii (Nogueira & Rodrigues, 2006), are probably part of this group.
Most studies mentioned above did not include data on Stenocercus dumerilii and S. tricristatus, this latter still known only from the type-specimen (Avila-Pires, 1995). Frost (1988) and Torres-Carvajal (2007a) examined for their studies a single cleared and double-stained skeleton of S. dumerilii, MCZ 160242. None of the two, however, presented a detailed description of the studied characters in the species. We present here a description of the cranium and abdominal skeleton of S. dumerilii, with emphasis on those characters used in the phylogenetic studies mentioned above.
MATERIALS AND METHODS
Two specimens of Stenocercus dumerilii of the herpetological collection of the Museu Paraense Emílio Goeldi, MPEG 7383 (<$, 63mm SVL) and MPEG 7332 (Ç, 95mm SVL), were cleared and stained following the methodology of Dingerkus & Uhler (1977). Description was mainly based on MPEG 7383, with MPEG 7332 used to verify some characters. The skull is described without separating the bones. A few internal skull structures (orbitosphenoid, epipterygoid), scleral ossicles, and cartilages are not described. Nomenclature followed Torres-Carvajal (2003, 2004) except when otherwise mentioned. Drawings were made with the aid of a stereomicroscope equipped with a camera lucida.
RESULTS
SKULL
The skull of Stenocercus dumerilii is amphikinetic, 24.5 mm long in MPEG 7332, 18.6 mm long in MPEG 7383; they are 89% as wide as long, and 51-53% as high as long. Along the middorsal line, the skull ascends gently up to the mid-posterior region of the frontal, then making a descending curve along the parietal. The frontal ascends moreover lateroposteriorly toward the postfrontal, which represents the highest point of the skull.
DERMATOCRANIUM
Premaxilla (Figures 2A-C, pm). Premaxillae fused into a single bone, forming the anteromedial margin of the snout and the medial margin of the fenestrae exonarinas. It has an acute triangular shape in dorsal view, with a broad, round-shaped anterior margin, and a posterior angular portion which extends between the nasals as a nasal process. On the alveolar portion there are two foramina, each about halfway between the mid-dorsal line and the lateral margin of the bone. In ventral view the premaxilla is separated from the vomers and bears eight monocuspid teeth anteroventrally and a posteromedial pair of incisive processes (Figure 2B, pip).
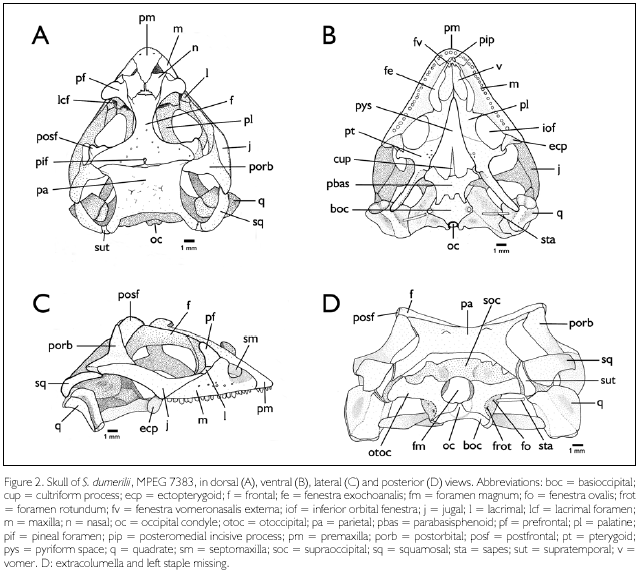
Septomaxillae (Figure 2C, sm). The septomaxillae, which partially separate at each side the nasal cavity and the cavum containing the vomeronasal organ, are compressed dorsoventrally, larger and with the sides curved upwards posteriorly.
Maxillae (Figures 2A-C, m). Each maxilla is subtriangular, with a concave surface anterodorsally corresponding to the ventral and lateroposterior walls of each fenestra exonarina. It is in contact with the premaxilla anteriorly, the septomaxilla anterodorsally, the nasal and prefrontal dorsally (on the nasal process), and the lacrimal and jugal posterodorsally. Twelve foramina are evident on each maxilla in MPEG 7383: one posteriorly on the concave surface; a row of seven foramina parallel to the ventral rim of the maxilla; three foramina approximately dorsal to the second foramen (anteroposteriorly) of this row; and another one dorsal to the third foramen. The posterior portion of the maxilla forms part of the floor of the orbit and it is dorsally overlapped by the prefrontal anteriorly, the jugal laterally, the ectopterygoid posteriorly and the palatine medially. Anterior to the inferior orbital fenestra the maxilla forms the floor of the maxillopalatine foramen, which is smaller than the lacrimal foramen. In ventral view the maxilla forms anteriorly the lateral margin of the fenestrae vomeronasalis externa and exochoanalis (Figure 2B, fv, fe), which are continuous, and posteriorly the lateral rim of the inferior orbital fenestrae (Figure 2B, iof). It presents three contact points: anteriorly with the premaxilla, where a medial process extends next to the ventroposterior margin of the premaxilla; medially with the palatine, through a triangular palatine process; and posteriorly with the ectopterigoyd bone, through a diagonal suture. In MPEG 7383 the right maxilla bears 18 teeth, the left one 17 teeth; in MPEG 7332 these numbers are 20 and 19, respectively. Teeth are pleurodont, compressed laterally and slightly recurved. Most of the teeth are tricuspid, bearing a medium cusp larger than the lateral ones, however all cuspids are small. The anterior maxillary teeth tend to be monocuspidal in shape.
Nasals (Figure 2A, n). Nasals flat, narrowly separated from each other medially. Anterior margin laterally (above septomaxilla) concave, medially covered by the premaxilla. Each nasal contacts laterally the nasal process of the maxilla, the prefrontal, and the anterolateral process of the frontal; posteriorly it partially overlaps the frontal. Between the two nasals and the frontal there is a non-ossified area (frontonasal fontanelle). Six foramina are evident on the dorsal surface of the left nasal and four on the right one (MPEG 7383).
Prefrontals (Figures 2A, 2C, pf). The two prefrontals form the anterodorsal rim of the orbits. Each bears three processes: (1) an anterodorsal nasal process partially separating the maxilla from the nasal bone; (2) a posterodorsal process forming a long suture with the frontal; and (3) a ventral process which contacts ventrally the palatine and ventrolaterally the lacrimal. The ventral process and the lacrimal delimit the lacrimal foramen (Figure 2A, lcf) and ventrally, together with the palatine and the maxilla, the maxillopalatine foramen; between these two foramina a space separates the ventral process of the prefrontal and the lacrimal. The central portion of the prefrontal forms a trihedral, recurved protuberance projected toward the orbital cavity.
Lacrimals (Figures 2A, 2C, l). Each lacrimal forms part of the orbit, between the jugal and the prefrontal; besides, anteriorly and ventrally it borders the maxilla. In lateral view the lacrimal has a subrectangular shape. In medial view its anterodorsal margin, which partially delimits the lacrimal foramen, is concave, and the anteroventral and ventral margins are convex, with irregular contours. Ventromedially it borders the maxillopalatine foramen.
Frontal (Figures 2A, 2C, f). The frontal forms most of the dorsal rim of the orbit. In dorsal view this bone is narrowest at midlength and distinctly wider posteriorly than anteriorly. An elongated, narrow, anterolateral process, at each side, partially separates the prefrontal from the nasal. The posterior processes are in contact with the postfrontal lateroanteriorly, the postorbital lateroposteriorly, and the parietal posteriorly. Medially in MPEG 7383 the posterior margin presents a short suture leading to the pineal foramen, which is completely enclosed in the frontal (Figure 2A, pif). The suture is slightly longer than the foramen. In MPEG 7332 the pineal foramen lies in the frontoparietal suture. Several small foramina occur radially at a certain distance from the pineal foramen.
Parietal (Figures 2A, 2D, pa). The parietal is nearly rectangular in shape (slightly wider than long), with two anterolateral processes in contact with the frontal and the postorbital. Each posterior corner extends posteriorly as a long supratemporal process, which overlaps dorsolaterally the supratemporal, and reaches medially the paraoccipital process of the otoccipital. Laterally the parietal forms the medial margin of the supratemporal fossa. In ventral aspect, at some distance from the border at each side, a lateral ridge runs from the suture with the frontal to the supratemporal process. Besides, the parietal is bound by connective tissue to the prootic posterolateraly, and to the supraoccipital posteriorly. The dorsal surface of the parietal presents a pair of low, conical protuberances positioned symmetrically at midlength, and a few foramina.
Supratemporals (Figures 2A, 2D, sut) The supratemporals are elongate and slender bones located between the parietal, squamosal, quadrate and the paraoccipital process of the otoccipital. Anteriorly it extends lateroventrally along the ventral ridge of the parietal, bordering the supratemporal fossa.
Postfrontals (Figures 2A, 2C, 2D, posf). Each postfrontal has a quadrangular and flat (in ventral view) base, and a concave, roughly triangular dorsolateral area. An angulate projection directed anteriorly into the orbit marks the dorsolateral deflection of the head, as well as its highest point. It is in contact with the frontal medially and the postorbital posteriorly. At least fifteen foramina are present on each postfrontal.
Postorbitals (Figures 2A, 2C, 2D, porb). Each postorbital is triradiate, with a dorsal process in contact with the postfrontal, frontal and parietal, which it partially overlaps; an anteroventral process and a posteroventral process, both of which border the posterodorsal margin of the jugal. The posterior process, together with the posterior extremity of the jugal, overlap the anterior end of the squamosal, with a flexible ligament separating them. The dorsal process of each postorbital forms the posteroventral rim of the orbit and the anterior rim of the supratemporal fossa. The posterior process and the squamosal form the supratemporal arch, delimiting the supratemporal fossa laterally. At least eleven foramina are present in each postorbital.
Squamosals (Figures 2A, 2C, 2D, sq). Each squamosal is elongate, pointed anteriorly, where it is overlapped by the postorbital and the jugal, and with a broader, round posterior end that reaches the supratemporal. The ventroposterior tip borders the cephalic condyle of the quadrate. It forms the posterior part of the supratemporal arch, which constitutes the lateral rim of the supratemporal fossa.
Jugals (Figures 2A-C, j). The jugals are elongate and depressed, with an anterior process dorsal to the posterior segment of the maxilla and reaching the lacrimal, and a posterior, wider process forming an angle of 130° upward in relation to the anterior process. The posterior process presents posterodorsally a long suture with the postorbital and its tip reaches shortly beyond the anterior tip ofthe squamosal, from which it is separated by a flexible ligament. Just anterior to the posterior process (at the posterior limit of the maxilla), the jugal articulates medially with the ectopterygoid. The jugals form the ventral rims of the orbit. Seven to eight foramina are present on their lateral surface.
Vomers (Figure 2B, v). The vomers are elongate and flat dorsoventrally, representing the most anterior elements of the palate. They meet each other medially for about two-thirds of their length, and are posteriorly separated by the vomerine processes of the palatines. A distinct groove is present at each vomer anteriorly, where it is separated from the premaxilla and the premaxillary process of the maxilla by connective tissue. The vomers form the medial borders of the fenestrae vomeronasales externae (Figure 2B, fv) and the anteromedial borders of the fenestrae exochoanales (Figure 2B, fe). Each vomer presents four or five foramina.
Palatines (Figures 2A-B, pl). Each palatine has three processes - a narrow vomerine process anteriorly, a short maxillary process laterally, and a wide and flat pterygoid process posteriorly. The vomerine process is in contact anteriorly, through a diagonal suture, with the vomers. The maxillary process presents a V-shaped suture with the maxilla. The suture with the pterygoid is also diagonal and hardly visible. The palatines form a medial suture at the most anterior part of the vomerine process; posteriorly they open into the pyriform space and extend to about half of its length (Figure 2B, pys). The lateral margin of the vomerine process and the anterior margin of the maxillary process of the palatine form the posterior border of the fenestra exochoanalis. The posterior margin of the maxillary process and the lateral margin of the pterygoid process form the anteromedial border of the inferior orbital fenestra (Figure 2B, iof). The maxillary process borders at its end the maxillopalatine foramen.
Ectopterygoids (Figures 2B-C, ecp). They are small and connect the pterygoid with the maxilla and the jugal. The lateral end of each ectopterygoid has two processes - an anterodorsal, pointed process that articulates with the posterior, dorsomedial end of the maxilla and the ventromedial surface of the jugal; and a ventroposterior, pointed process that borders the medial posterior margin of the maxilla and the medial surface of the jugal, close to the anterior portion of its ascending segment. Medially, each ectopterygoid bifurcates into an anterodorsal and a ventral digit-like extensions that brace the transverse process of the pterygoid. The ectopterygoids form the posterolateral border of the inferior orbital fenestra (Figure 2B, iof).
Pterygoids (Figure 2B, pt). The pterygoids are long, each with three processes: the anterior, mostly laminar palatine process; the lateral, short and diagonally flattened transverse process; and the posterior, depressed quadrate process. The palatine process is continuous with the pterygoid process of the palatine, the suture between them being hardly visible. Posteromedially it tickens and bears 1-4 teeth of variable sizes. The transverse process tickens into a knob distally, which is braced by the ectopterygoid. Medially, at the opposite side of the transverse process, the pterygoid presents a vertical concavity and a dorsal projection, which articulates with the basipterygoid process of the parabasisphenoid. The quadrate process reaches the medial posterior portion of the quadrate; its medial surface forms a shallow concavity. The anterior end of the quadrate process bears dorsally the columellar fossa and, a short distance posterior to it, a small, conical elevation (postcolumellar process of Torres-Carvajal, 2003). The pterygoids form the posteromedial rim of the inferior orbital fenestrae (Figure 2B, iof) and the posterior lateral margins of the pyriform space (Figure 2B, pys).
Dentaries (Figure 3A-B, d). The dentaries form the anterior margin of the mandible. In labial aspect it represents about 60% of the total length of the mandible, reaching beyond the level of the coronoid apex, but not beyond the level of the coronoid posterior process. The alveolar sulci are well differentiated and they bear 22-23 teeth in MPEG 7383, 26 teeth in MPEG 7332, of which the anterior one to three are monocuspid, the posterior ones tricuspid. Meckel's canal is fused throughout most of its length and exposed between the first and sixth teeth (Figure 3A, mc). In lingual aspect the posterior margin of each dentary bifurcates and overlaps the splenial, to articulate with the coronoid dorsally and the angular ventrally. In labial aspect, each dentary articulates with the labial process ofthe coronoid, the supra-angular and the angular. The dorsal border of the dentary at the contact point with the coronoid is above the level of the dorsal border of the supra-angular. Labially each dentary bears five foramina aligned longitudinally on its anterior third.
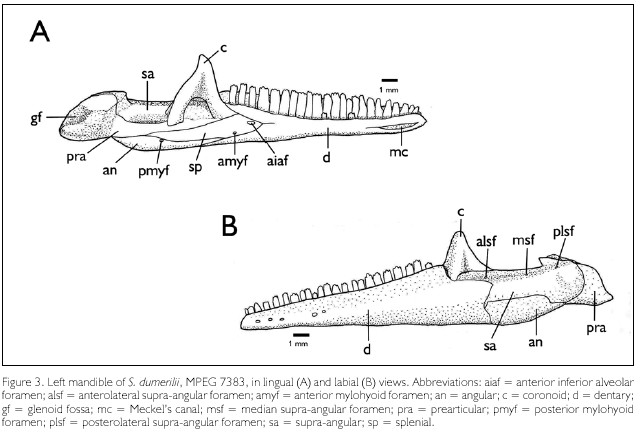
Coronoids (Figures 3A-B, c). Each coronoid projects dorsally posterior to the dentary teeth, its height approximately similar to that of the dentary. This dorsal projection is triangular and laterally flattened. Ventrally one labial and two lingual processes overlap the supra-angular. The labial process borders anteriorly the posterodorsal margin of the dentary; it reaches about the level (dorso-ventrally) of the supra-angular anterolateral foramen, and hardly overlaps the dentary. Besides the supra-angular, the lingual anterior process of the coronoid is in contact with the dentary anteriorly, the splenial ventrally, and the prearticular posteroventrally. The lingual posterior process overlaps the prearticular ventral to the supra-angular.
Supra-angulars (Figures 3A-B, sa). Each supra-angular is about half the maximal length of the dentary. The supra-angular forms the posterodorsal segment of the mandible, with a distinct dorsal horizontal surface. In dorsolateral view the anterior margin of the supra-angular bifurcates at the contact with the dentary, forming a long dorsal process, which reaches the labial process of the coronoid, and a short ventral process inserted between the dentary and the angular. At about two-thirds of its length the ventral margin of the supra-angular makes a round curve downward, following the narrowing of the angular, which it shortly overlaps, and then an angle turning again posteriorly until it reaches the prearticular. In lingual aspect the supra-angular is overlapped by the coronoid anterodorsally, but it is visible between, and posterior to, the two lingual processes of the coronoid. It is in contact ventrally with the prearticular, but posterior to the coronoid the two bones are partially separated by the adductor fossa. Posteriorly the supra-angular is in contact with the prearticular and articular. Dorsomedially it presents a low protuberance close to its posterior border, and the anterodorsal border rises into a pointed tip. Three foramina occur on the dorsal surface of each supra-angular - the supra-angular anterolateral foramen (Figure 3B, alsf), on the anterolateral dorsal process; the supra-angular posterolateral foramen (Figure 3B, plsf), smaller than the former, at the base of the dorsomedial protuberance mentioned above; and the median supra-angular foramen (Figure 3B, msf), smaller than the previous two and located halfway between them.
Prearticulars (Figures 3A-B, pra). Only the posterior part of each prearticular is seen in labial aspect. In lingual aspect, an elongate anterior process reaches the posterior margin of the lingual anterior process of the coronoid. The prearticular is dorsally in contact with the supra-angular, is overlaid by the lingual posterior process of the coronoid, and forms the ventral margin of the adductor fossa. Ventrally it borders the splenial and the angular. The posterior segment - the prearticular process -, together with the articular (the limits between these two elements are not clear), presents a concave ventral surface, a concave posterodorsal surface, and a concave dorsal surface, the latter two forming the glenoid fossa (Figure 3A, gf). There is no angular process.
Angulars (Figures 3A-B, an). They form the ventroposterior surface of the mandible. In labial aspect each angular has the same height for about three fourths of its length, but posteriorly it narrows into a pointed end. It contacts the dentary anteriorly, the supra-angular dorsally, and the prearticular posteriorly. In lingual aspect a sharply pointed anterior process is exposed between the splenial and the dentary, and posteriorly it borders the prearticular. Close to the posterior end of the suture with the splenial lies the posterior mylohyoid foramen (Figure 3A, pmyf).
Splenials (Figure 3A, sp). The splenials are only seen in lingual aspect. Anteriorly, they are overlapped by the bifurcation of the posterior margin of the dentaries. Posteriorly, each splenial is in contact with the anterior lingual process of the coronoid and the prearticular dorsally, and with the angular ventrally. The large anterior inferior alveolar foramen (Figure 3A, aiaf) lies next to the margin of the dorsal segment of the dentary. Ventrally, shortly anterior to the border with the angular, lies the anterior mylohyoid foramen (Figure 3A, amyf), smaller than the anterior inferior alveolar foramen. Besides, two other foramina are present, posterior to, and about in line with, the alveolar foramen; the anterior of the two is about similar in size to the anterior mylohyoid foramen, the posterior one distinctly smaller.NEUROCRANIUM
Basioccipital (Figures 2B, 2D, boc). The basioccipital forms the posterior floor of the braincase, with a wide, interdigitate suture with the parabasisphenoid anteriorly - two short anterior processes of the basioccipital lies between two posterolateral longer processes and a posteromedial short process of the parabasisphenoid (the parabasisphenoid processes partially overlapping the basioccipital). Posteriorly it forms the medial portion of the occipital condyle (Figures 2B, D, oc), and laterally it bears on each side the ventrally projected spheno-occipital tubercle. At each side the basioccipital contacts anterodorsally the inferior process of the prootic, and posterodorsally the crista interfenestralis and the crista tuberalis of the otoccipital. The spheno-occipital tubercle lies between these two cristae and their walls form the antrum of the foramen rotundum (Figure 2D, frot).
Supraoccipital (Figure 2D, soc). A saddle-like bone, with an elevated middorsal area that becomes wider anteriorly, and posteriorly to a lesser degree; at both sides of this elevated middorsal area there is a shallow depression. The supraoccipital lies ventroposteriorly of the parietal, from which it is separated by connective tissue, except for the processus ascendens, a thin anteromedial extension of about one third the length of the parietal. The supraoccipital articulates anterolaterally with the alar process of the prootic, and posterolaterally with the otoccipital. It forms the posterodorsal wall of the braincase, takes part in the auditory bulla, and its ventroposterior margin forms the dorsal rim of the foramen magnum (Figure 2D, fm).
Parabasisphenoid (Figure 2B, pbas). This element corresponds to the fusion of the parasphenoid and the basisphenoid. Its central, slightly concave area forms the anterior floor of the braincase. It bears a long and thin anterior cultriform process (Figure 2B, cup) that extends anteriorly for more than half the length of the pyriform space, and in which only the base is ossified; two robust anterolateral basipterygoid processes, which articulate on each side with the meniscus pterygoideus of the pterygoid; two lateroposterior and one short midposterior processes that interdigitate with, and partially overlap (in ventral view), the basioccipital. Laterally, a dorsal extension is present at the base of each anterolateral process, and between these two surfaces is the entrance of a canal (corresponding to the Vidian canal mentioned by Torres-Carvajal, 2003). Each dorsal extension is in contact posteriorly with the prootic.
Otoccipitals (Figure 2D, otoc). Each otoccipital corresponds to the fusion of the opisthotic and the exoccipital at each side of the foramen magnum. They form the posterior wall of the braincase. The medial portion of each otoccipital is globose, forming part (together with the supraoccipital) of the auditory bulla. Dorsally it presents a wide suture with the supraoccipital, medially it forms the lateral margin of the foramen magnum and the lateral portion of the occipital condyle, and ventrally it contacts widely the basioccipital. Laterally each otoccipital has a paraoccipital process which articulates distally with the distal end of the supratemporal process of the parietal and the supratemporal bone. Anterolaterally each otoccipital contacts the prootic. At the border of the otoccipital and the prootic lies the fenestra ovalis, which is directed laterally (Figure 2D, fo) and receives the stapes. Ventromedially to the fenestra ovalis, between the crista interfenestralis and the crista tuberalis of the otoccipital, and (shortly) the spheno-occipital tubercle of the basioccipital, lies the foramen rotundum. Close to the rim of the occipital condyle lies the vagal foramen.
Prootics. Each prootic is a triradiate bone, with an alar process (directed anterodorsally), an anteroventral process, and a posterodorsal process. The alar and the anteroventral processes form the anterior portion of the lateral wall of the braincase. The posterior portion of the prootics forms part of the internal ear at each side. Each prootic contacts the supraoccipital through the alar process; the paraoccipital process of the otoccipital through the posterodorsal process; and the basioccipital and the parabasisphenoid through the anteroventral process. Between the prootic and the otoccipital lies the fenestra ovalis (Figure 2D, fo). The posterodorsal process is shortly overlapped laterally by the dorsomedial border of the quadrate, from which it is separated by connective tissue.
SPLANCHNOCRANIUM
Quadrates (Figures 2A-D, q). The quadrates articulate the skull with the mandible through their ventral, double condyle. Each quadrate is higher than wide, and it is bowed forward. The anterior aspect forms a wide, slightly concave surface. Posteriorly it presents a prominent vertical crest uniting the ventral condyle with a cephalic condyle that articulates with the supratemporal. The segment lateral to the vertical crest is wider than the medial segment, concave and bordered by the tympanic crest; it presents a short notch dorsolaterally, the superior fossa. The medial segment is wider and slightly concave dorsally, where it receives the processus internus of the extracollumela, and bordered by the medial crest. Dorsolaterally the quadrate articulates with the ventroposterior tip of the squamosal, dorsomedially with the posterodorsal process of the prootic, and medially, shortly dorsal to the ventral condyle, it receives the quadrate process of the pterygoid.
Stapes (Figures 2B, 2D, sta). The stapes are rod-shaped and form with the cartilaginous extracolumella the middle ear. It extends from the fenestra ovalis, between the prootic and the otoccipital, to about the midlevel of the medial segment of the quadrate, with which it attaches through the extracolumella.
ABDOMINAL SKELETON
Sternum, sternal ribs, and inscriptional ribs (Figures 4A-B). The sternum presents a single median fontanelle, which lies shortly posterior to, and is slightly wider than, the posterior process of the interclavicle. Three pairs of sternal ribs. Xiphisternal rods elongate and ending in a small, recurved segment; it gives rise to a single pair of inscriptional ribs. Five pairs of short postxiphisternal inscriptional ribs; none articulates with its pair, or approximates the midventral line. Both MPEG 7332 and MPEG 7383 present the same numbers of inscriptional ribs.
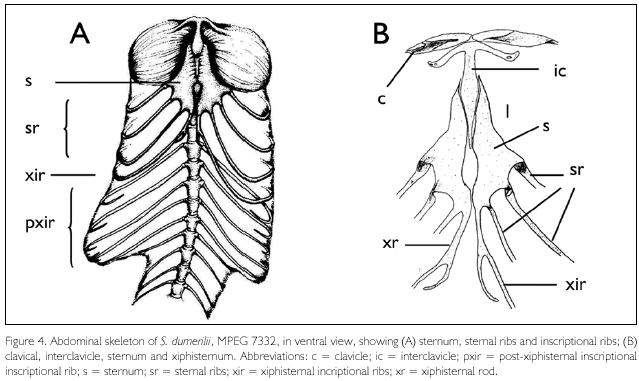
Interclavicle (Figure 4B). The interclavicle is arrow-shaped, with the lateral processes forming an angle of about 50°-60° with the posterior process. The posterior process comes close to, but does not reach, the level of the lateral corners of the sternum, which approximately coincides with the anterior margin of the sternal fontanelle. The free segment (not in contact with the sternum) of the posterior process of the interclavicle corresponds to about 0.35-0.4 times the length of the sternum.
Clavicles (Figure 4B). The clavicles are elongate, recurved, with a flat posterior expansion medially, for about half of its length; this expansion in only lightly ossified in MPEG 7332, more so in MPEG 7383. No foramen or ornamentation is present.
The only Stenocercus species of which an extensive description of the cranium is available is S. guentheri (Boulenger, 1885) (Torres-Carvajal, 2003). Shape of the head in S. dumerilii is quite distinct from that in most Stenocercus species, including S. guentheri, because of the sharply demarcated canthal-supraciliary crest, ending in an enlarged, horn-like scale (Figure 1). Some differences found between the two species result most probably from that. Most obvious ones are the shape of the prefrontals and postfrontals, more prominent in S. dumerilii. Maybe also related is the more pronounced widening of the frontal posteriorly, touching the postorbital, in S. dumerilii, while in S. guentheri it is separated from the postorbital by the parietal. Still in this general region of the skull, S. guentheri presents an anterodorsal process on the postorbital, not found in S. dumerilii. Other differences are also found. The premaxilla differs in shape (especially its nasal process) in both species, but they are similar in bearing ventrally posteromedial incisive processes and in not presenting posteroventral extensions at their articulation with the maxilla. The infraobrital foramen is formed in S. dumerilii by the prefrontal and the palatine, while in S. guentheri the maxilla is also involved. The pineal foramen is completely enclosed by the frontal in one of the specimens of S. dumerilii (MPEG 7383), and between the frontal and the parietal in the other (MPEG 7332); in S. guentheri it was observed always between the frontal and the parietal, even though with variation on how much each bone contributed to it. Concerning the mandibular bones, the main difference is the absence of the prearticular angular process in S. dumerilii, a character variable in other groups of lizards as well and that may show ontogenetic variation, becoming relatively larger as the animal grows (e.g., in most iguanines, Queiroz, 1987). Moreover, the dentary in S. guentheri extends in labial aspect much posterior to the coronoids, while in S. dumerilii it reaches approximately the posterior level of the coronoids. The anterior end of the Meckel's canal also differs, opened only ventral to the second tooth in S. guentheri, between the first and sixth teeth in S. dumerilii.
Cadle (1991) described some osteological characters from four species of Stenocercus. In all of them he observed that the region of the nasal-frontal contact was well ossified. Torres-Carvajal (2003) observed that in individuals smaller than 39 mm SVL in S. guentheri the frontonasal fontanelle (at the nasal-frontal contact) was still present, disappearing in larger specimens. In S. dumerilii the specimens studied had SVL of 63 mm and 95 mm and the nasal-frontal region was not yet completely ossified.
The following characters in S. dumerilii agree with its taxonomic position within tropidurids and more inclusive taxa, according to the studies by Frost & Etheridge (1989), Frost (1992), and Torres-Carvajal (2007b): (1) premaxilla overlaps nasals; (2) maxillae not meeting anteromedially posterior to palatal portion of premaxilla; (3) skull roof not strongly rugose; (4) jugal and squamosal not broadly juxtaposed; (5) parietal roof trapezoidal in shape; (6) supratemporal sits mostly on lateral side of supratemporal process of parietal; (7) superior fossa of quadrate not enlarged; (8) dentary not expanded onto labial face of coronoid; (9) labial blade of the coronoid poorly developed; (10) anterolateral supra-angular foramen dorsal to posterior extremity of the dentary; (11) Meckel's groove partially fused; (12) splenial very short anteriorly (less than 1/6 length tooth row); (13) pleurodont teeth, not fused to underlying bone; (14) palatine teeth absent; (15) pterygoid teeth present; (16) anterior process of interclavicle absent; (17) posterior process of interclavicle not invested by sternum far anteriorly; (18) sternal fontanelle median and enlarged.
Frost & Etheridge (1989) mentioned as a character of Tropiduridae the "lacrimal foramen enlarged" when compared to the maxillopalatine foramen. These two foramina in S. dumerilii are similar to those described by Torres-Carvajal (2003) in S. guentheri (except that in S. dumerilii the lacrimal reaches the border of the maxillopalatine foramen) - in both species the lacrimal foramen is distinctly larger than the maxillopalatine foramen. Frost (1992) also reported within the "Tropidurus group" that the maxillopalatine foramen may be much smaller than, or subequal in size to, the lacrimal foramen. Therefore the above character mentioned by Frost & Etheridge (1989) for Tropiduridae is not valid. Also considered a character of Tropiduridae by these authors was the position of the pineal foramen in the frontoparietal suture (or absent, as opposed to "entirely within the frontal"). This is in agreement to what was observed in S. guentheri by Torres-Carvajal (2003) and in one of the specimens of S. dumerilii studied (MPEG 7332), but not in the other (MPEG 7383), where the pineal foramen is within the frontal, thus showing variation in this character in the species.
As observed by Frost (1992) and Torres-Carvajal (2007a), the postxiphisternal inscriptional ribs (PIR) are not in contact midventrally (a plesiomorphy in Stenocercus). A midventral contact, or close proximity, between at least one pair of PIR was proposed as diagnostic of "Ophryoessoides" sensu Etheridge (1966) and sensu Fritts (1974), the nominal taxon where this species was originally described. On the other hand, S. dumerilii seems to be unique in its pattern of abdominal rib attachment, with one xiphisternal (XIR) and five postxiphisternal pairs of inscriptional ribs; the xiphisternal longitudinal rods extend posterior to the articulation with the XIR, forming posterior xiphisternal processes. A single pair of xiphisternal ribs has been reported within tropidurids only in Uracentron azureum (Etheridge, 1959). Torres-Carvajal (2004, 2007a) found in all tropidurid specimens he examined 2-3 XIR and 2-4 PIR. Among his material was a specimen of U. azureum, thus indicating intraspecific variation in this character. Posterior xiphisternal processes also have been observed in some other species, as well as intraspecific variation in their presence and shape (Torres-Carvajal, 2004). It is not possible to rule out, therefore, that the pattern of abdominal rib attachment in S. dumerilii is more variable than described here, and that at least part of the specimens present also two XIR, as it is most commonly found in tropidurids. On the other hand, this character has not been studied in those species considered most closely allied to S. dumerilii (S. tricristatus, S. quinarius and S. squarrosus), leaving also the possibility that they share this pattern.
The three above-mentioned species have also in common with S. dumerilii a pyramidal head with a distinct canthal-superciliary ridge that ends in a prominent postsupraciliary scale (Nogueira & Rodrigues, 2006). As noted above, these characters in S. dumerilii are also reflected in the cranium, especially in the shape ofthe prefrontal bone, which bears a trihedral, recurved protuberance projecting towards the orbital cavity, and in the shape of the postfrontal bone, which has an angulate projection directed anteriorly into the orbital cavity. The postfrontal represents the highest point of the cranium. The study of the crania in the three species considered most closely allied to S. dumerilii would help to indicate whether the shape ofthe head is homologous in these taxa, which would corroborate monophyly of this group.
ACKNOWLEDGEMENTS
We thank the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Brazil, for a master scholarship to EJHR; Ana Lúcia da Costa Prudente, Miguel Trefaut Rodrigues and Maria Luiza Videira Marceliano for suggestions to improve an earlier manuscript on this subject that served as basis for this paper; and Marcelo José Sturaro for the final composition of figures 2 and 3. EJHRthanks Carlos Prada for helping with the English translation of a preliminary version of this paper.
REFERENCES
ANDRADE, G., J. D. LIMA & A. O. MACIEL, 2003. Geographic Distribution. Stenocercus dumerilii. Herpetological Review 34(4): 385-386.
AVILA-PIRES, T. C. S., 1995. Lizards of Brazilian Amazonia (Reptilia: Squamata). Zoologische Verhandelingen Leiden 299: 1-706.
BOULENGER, G. A., 1885. Catalogue ofthe lizards in the British Museum (Natural History). Trustees of the British Museum 2: 1-497.
CADLE, J. E., 2001. A new species of lizard related to Stenocercus caducus (Cope) (Squamata: Iguanidae) from Peru and Bolivia, with a key to the "Ophryoessoides group". Bulletin of the Museum of Comparative Zoology 157(3): 183-221.
CADLE, J. E., 1998. New species of lizards, genus Stenocercus (Iguania: Tropiduridae), from western Ecuador and Peru. Bulletin of the Museum of Comparative Zoology 155(6): 257-297.
CADLE, J. E., 1991. Systematics of lizards of the genus Stenocercus (Iguania: Tropiduridae) from northern Peru: new species and comments on relationships and distribution patterns. Proceedings of the Academy of Natural Sciences of Philadelphia 143: 1-96.
CUNHA, O. R., 1981. Lacertílios da Amazônia VIII - Sobre Ophryessoides tricristatus Dumeril, 1851, com redescrição da espécie e notas sobre ecologia e distribuição na região leste do Pará (Lacertilia, Iguanidae). Boletim do Museu Paraense Emílio Goeldi, série Zoologia 108: 1-23.
DINGERKUS, G. & L. D. UHLER, 1977. Enzyme clearing of alcian blue stained whole small vertebrates for demostration of cartilage. Stain Technology 52(4): 229-232.
ETHERIDGE, R. E., 1966. The systematic relationships of West Indian and South American lizards refered to the iguanid genus Leiocephalus. Copeia 1: 79-91.
ETHERIDGE, R. E., 1959. The relationships of the anoles (Reptilia: Sauria: Iguanidae). An interpretation based on skeletal morphology: 1-236. Ph.D. Dissertation - University of Michigan, Ann Arbor, Michigan.
FRITTS, T. H., 1974. A multivariate evolutionary analysis of the Andean Iguanid lizards of the genus Stenocercus. Memoir San Diego Society of Natural History 7: 1-89.
FROST, D. R., 1992. Phylogenetic analysis and taxonomy of the Tropidurus group lizards (Iguania: Tropiduridae). American Museum Novitates 3033: 1-68.
FROST, D. R., 1988. A phylogenetic analysis of the Tropidurus group of iguanian lizards, with comments on the relationships within the Iguania (Squamata): 1-350. Ph.D. Dissertation -University of Kansas, Lawrence.
FROST, D. R. & R. ETHERIDGE, 1989. A phylogenetic analysis and taxonomy of Iguanian lizards (Reptilia: Squamata). Miscellaneous Publications the University of Kansas Museum of Natural History 81: 1-65.
FROST, D. R., R. ETHERIDGE, D. JANIES & T. A. TITUS, 2001. Total evidence, sequence alignment, evolution of Polychrotid lizards, and a reclassification of the Iguania (Squamata: Iguania). American Museum Novitates 3343: 1-38.
NOGUEIRA, C. & M. T. RODRIGUES, 2006. The genus Stenocercus (Squamata: Tropiduridae) in extra-Amazonian Brazil, with the description of two new species. South American Journal of Herpetology 1(3): 149-165.
QUEIROZ, K., 1987. Phylogenetic systematics of iguanine lizards. A comparative osteological study. University of California Publications in Zoology 118: 1-203.
SCHULTE, J. A., J. P. VALLADARES & A. LARSON, 2003. Phylogenetic relationships within Iguanidae inferred using molecular and morphological data and a phylogenetic taxonomy of iguanian lizards. Herpetologica 59(3): 399-419.
SEMA-PA, 2007. Resolução COEMA no 54, de 24 de outubro de 2007. Homologa a lista de espécies da flora e da fauna ameaçadas no Estado do Pará. Available in: <http://www.sema.pa.gov.br/resolucoes_detalhes. php?idresolucao=54>. Access on: 06 June 2008.
SILVA, J. M. C., A. B. RYLANDS & G. A. B. FONSECA, 2005. The fate of the Amazonian areas of endemism. Conservation Biology 19(3): 689-694.
STEINDACHNER, F., 1867. Reise der österreichischen Fregatte Novara um die Erde in de Jahren 1857, 1858, 1859 unter den Benfehlen des commodore B. von Wüllerstorf-Urbair: 1-98. Zoologischer Theil, Reptilien, Wien.
TORRES-CARVAJAL, O., 2007a. Phylogeny and biogeography of a large radiation of Andean lizards (Iguania, Stenocercus). Zoologica Scripta 36(4): 311-326.
TORRES-CARVAJAL, O., 2007b. A taxonomic revision of South American Stenocercus (Squamata: Iguania) lizards. Herpetological Monographs 21: 76-178.
TORRES-CARVAJAL, O., 2004. The abdominal skeleton of Tropidurid lizards (Squamata: Tropiduridae). Herpetologica 60(1): 75-83.
TORRES-CARVAJAL, O., 2003. Cranial osteology ofthe andean lizard Stenocercus guentheri (Squamata: Tropiduridae) and its postembryonic development. Journal of Morphology 255(1): 94-113.
TORRES-CARVAJAL, O., J. A. SCHULTE & J. E. CADLE, 2006. Phylogenetic relationships of South American lizards of the genus Stenocercus (Squamata: Iguania): a new approach using a general mixture model for gene sequence data. Molecular Phylogenetics and Evolution 39: 171-185.
 Mailing address:
Mailing address:
Museu Paraense Emílio Goeldi
Editor do Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais
Av. Magalhães Barata, 376
São Braz – CEP 66040-170
Belém - PA - Brazil
Caixa Postal 399
Phone: 55-91-3182-3246
Fax: 55- 91-3249-6373
E-mail: boletim.naturais@museu-goeldi.br
Recebido: 21/08/2008
Aprovado: 05/12/2008
