










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.1 n.2 Belém ago. 2006
Relações da biomassa microbiana do solo com características químicas de frações orgânicas e minerais do solo após exploração mineral na Amazônia Oriental1
Relationships of soil microbial biomass with the chemical characteristics of organic and mineral soil fractions after mineral exploration in Eastern Brazilian Amazon
Maria de Lourdes Pinheiro RuivoI; Nairam Félix de BarrosII; Carlos Ernesto Gonçalves Reynaud SchaeferIII
IMuseu Paraense Emílio Goeldi. Coordenação de Ciências da Terra e Ecologia. Belém, Pará, Brasil (ruivo@museu-goeldi.br)
IIUniversidade Federal de Viçosa. Departamento de Solos. Viçosa, Minas Gerais, Brasil (nfbarros@ufv.br)
IIIUniversidade Federal de Viçosa. Departamento de Solos. Bolsista do CNPq. Viçosa, Minas Gerais, Brasil (carlos.schaefer@ufv.br)
RESUMO
Foram avaliadas as características químicas e biológicas de solos alterados da mina de bauxita de Porto Trombetas, município de Oriximiná, no estado do Pará, Brasil a partir de sítios recuperados entre 1981 e 1994 e do solo inalterado (latossolo amarelo). Os valores referentes às características químicas e biológicas foram obtidos a partir das análises da manta orgânica, matéria orgânica leve e do solo, dos sítios inalterado e alterado. Os dados foram submetidos a análises de correlação de Pearson e de Trilha. Os resultados mostram que a composição química da manta orgânica e da matéria orgânica leve relacionaram-se favoravelmente com os teores de C e P da biomassa microbiana do solo.
Palavras-chave: Biomassa microbiana do solo. Solos alterados. Mineração. Amazônia Oriental.
ABSTRACT
The studied area is located in Porto Trombetas, district of Oriximiná, in the northwest of Pará, state, Brazil and has been subject to bauxite mining. The chemical and biological measurements were made on sites recuperation from 1981 to 1994, and one in Yellow Latosol under secondary forest, taken as reference. The values corresponding to the chemical and biological characteristics were obtained from analyses of the litter layer, light organic matter and soil of the altered and unaltered sites. The data were analyzed using Pearson correlation and Path Analysis. The results given how the chemical an biological composition litter layer and light organic matter were related of the carbon and microbial biomass phosphorus concentration.
Keywords: Soil microbial biomass. Altered soils. Mining. Western Amazon.
INTRODUÇÃO
A revegetação de terrenos degradados é, em parte, dependente do retorno do horizonte A à área revegetada e da colonização da população microbiana no solo após a lavra, que favorecerá o incremento da atividade microbiana (TRINDADE; TÓTOLA; DIAS, 1995). Assim, a interrelação entre desenvolvimento vegetal e atividade microbiana é um fato importante na recuperação de solos alterados pela mineração.
Durante a recuperação paisagística, a comunidade microbiana é constantemente mudada (TURCO; KENNEDY; JAWSON, 1994). Solos exauridos são comumente caracterizados por população microbiana muito baixa, menor variedade de espécies e gêneros de bactérias e baixa taxa de decomposição da celulose, quando comparados com os solos inalterados (SEGAL; MANCINELLI, 1987).
A cobertura vegetal, além de favorecer a atividade de organismos, é responsável pelo aporte de matéria orgânica ao solo que, por meio de sua mineralização, concorre para maior disponibilidade de nutrientes para as plantas. A manta orgânica é o principal componente de retorno de C ao solo, sendo, portanto, de grande relevância na ciclagem de nutrientes e de carbono. Sua taxa de decomposição depende, entre outros fatores, da qualidade do material, sendo o teor de lignina um aspecto importante.
Nos ecossistemas florestais com relação C/N estreita, a decomposição da manta é rápida. Para um dado sítio, porém, a taxa de decomposição pode estar relacionada com a atividade e com a composição dos organismos do solo. Oliveira (1997), analisando a diversidade de grupos taxonômicos de mesofauna em áreas recuperadas após lavra de bauxita em Porto Trombetas, verificou que nos plantios da década de 1980, considerados já bem estabelecidos, grupos-chave da decomposição da manta orgânica, como Isopoda sp. e Dipoplopoda sp, estavam mal representados, o que dificultou a decomposição deste material. No entanto, tanto a diversidade vegetal quanto a produção da manta orgânica foram importantes para a colonização e manutenção dos invertebrados terrestres.
Duda et al. (1999), ao realizarem estudos envolvendo a avaliação da dinâmica da matéria orgânica do solo para a caracterização de áreas degradadas, mostraram que a matéria orgânica leve do solo (MOL) pode ser usada como indicador da sua degradação ou recuperação. A MOL constitui-se, principalmente, de partes de plantas, embora resíduos de animais e de microrganismos também possam estar presentes em vários estágios de decomposição (KÖGEL-KNABNER; ZIEGLER, 1993). A quantidade de MOL é bastante influenciada pelo clima, características do solo, atividade da mesofauna, uso do solo e tipo de vegetação, entre outros fatores (CHRISTENSEN, 1992).
Este trabalho teve como objetivo principal avaliar a relação da biomassa microbiana do solo com as características químicas da manta orgânica, da matéria orgânica leve e do solo.
MATERIAL E MÉTODOS
A amostragem foi feita em julho de 1996 na mina de Bauxita do platô Saracá, na localidade de Porto Trombetas, município de Oriximiná, Pará, pertencente à Companhia de Mineração Rio do Norte (MRN). Foram selecionados para estudo 14 sítios, recuperados entre 1981 e 1994, e um local não-minerado em latossolo amarelo (LA) sob floresta densa perturbada. Algumas características dos sítios e de seus substratos estão listadas na Tabela 1.
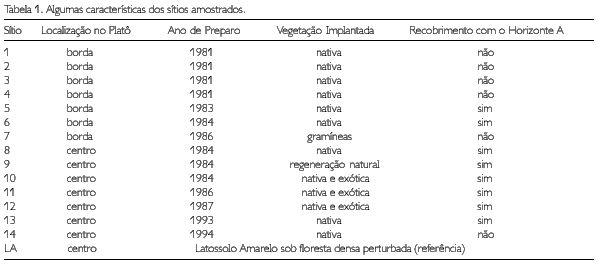
As áreas apresentam grande heterogeneidade quanto às características dos substratos, dificultando a caracterização. Então, para realizar o recobrimento vegetal, foram utilizadas espécies florestais nativas a partir da coleta de sementes na floresta e do preparo de mudas em viveiros, e espécies exóticas, à exceção do sítio 9, onde a cobertura vegetal se desenvolveu a partir de regeneração natural, e do sítio 7, onde foram plantadas exclusivamente gramíneas. Em todos os sítios, as plantas receberam 50 g por cova da mistura NPK 12-36-12. O sítio 10 (plantio experimental de Terminalia cattapa, com posterior regeneração natural da vegetação), ao contrário dos demais, recebeu 50 g por cova de superfosfato simples. Maiores detalhes acerca da lavra e recuperação da cobertura vegetal são encontrados em Ruivo (1991).
Para avaliação da biomassa microbiana, amostras compostas de solo, constituídas de dez amostras simples, foram coletadas ao acaso à profundidade de 0 a 2,5 cm, preservadas à temperatura de 4oC e enviadas ao laboratório. Neste, as amostras foram peneiradas em peneiras de 2 mm de malha e reumedecidas para atingirem 40% da capacidade de campo. A estimativa dos teores de carbono (CBM), nitrogênio (NBM) e fósforo (PBM) da biomassa microbiana foi feita utilizando-se o método da fumigação-extração (VANCE et al., 1987; GUERRA, 1993).
Para determinação da CBM e do NBM, pesaram-se 25 g de solo, em triplicata, fumigando-os por 24 h, em dessecador, com 2 ml de clorofórmio. Amostras não-fumigadas e fumigadas foram agitadas, por 30 min, com 50 ml de solução K2SO4 0,5 mol/l. No extrato, o CBM foi determinado pela digestão com dicromato de potássio e o nitrogênio total pelo método Kjeldahl. Para cálculo do CBM e NBM, os teores de C e N das amostras não-fumigadas foram subtraídos dos valores das amostras fumigadas, sendo a diferença dividida pelo valor de Kn = 0,54 (BROOKS et al., 1985), para obtenção do NBM, e de Kc = 0,26 (FEIGL et al., 1995), para obtenção do CBM.
Na determinação do PBM, também analisado com amostras em triplicata, foram usados 10 g de solo, fumigados por 24 h e colocados dentro do frasco 1 ml de clorofórmio. Amostras fumigadas e não-fumigadas foram agitadas por 30 min com 100 ml de solução NaHCO3 0,5 mol/l (pH 8,5). A estimativa do PBM foi feita a partir da determinação do P no extrato, após a formação da cor azul do complexo fosfato-molibdato em meio ácido (ácido sulfúrico) na presença de ácido ascórbico (EMBRAPA, 1997). A dosagem de P foi feita em espectrofotômetro, utilizando-se comprimento de onda de 660 nm. Para cálculo do PBM, os valores obtidos das amostras não-fumigadas foram subtraídos dos valores das amostras fumigadas, sendo a diferença dividida pelo valor de Kp = 0,40 (BROOKS; POWLSON; JENKINSON, 1982).
A manta orgânica (MOG) foi coletada em parcela de 0,3 x 0,3 m, também próxima dos locais onde foi amostrado o solo, para determinação da biomassa microbiana. Cada amostra foi composta por cinco amostras simples. A MOG foi seca em estufa a 68oC, por 48 h, pesada e triturada. Para determinação da matéria orgânica leve (MOL), foram coletadas amostras compostas de solo à profundidade de 0 a 2,5 cm, constituídas de cinco amostras simples, coletadas ao acaso. Tomaram-se 250 g de amostra de solo, da qual foram separadas a parte mineral e a parte orgânica por flotação em água com o auxílio de peneira de 0,25 mm. Após esta fase, a MOL foi seca em estufa a 68oC por 42 h; em seguida, o material foi pesado e triturado.
Tanto a MOG como a MOL, após a trituração, foram extraídas por meio de digestão nitro-perclórica e sulfúrica, para determinação dos cátions totais (Ca2+ e Mg2+, por espectrofotometria de absorção atômica, e K+, por fotometria de chama), do fósforo total (colorimetria) e do nitrogênio total (Kjeldahl), respectivamente. Foram também determinados os teores de lignina (Lg), celulose (Ce) e hemicelulose (He), segundo Soest (1963).
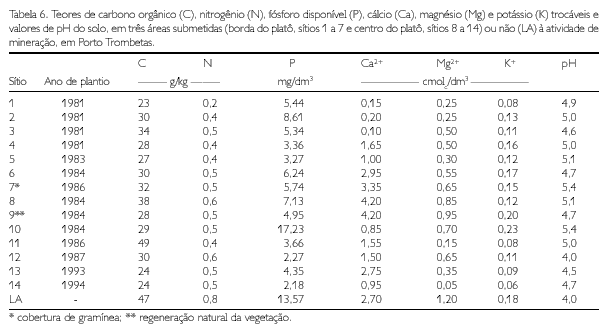
Em cada um dos sítios selecionados para o estudo, foram coletadas amostras compostas de solo, a partir de cinco amostras simples, na profundidade de 0 a 2,5 cm, nas proximidades dos locais onde foi amostrado o solo para determinação da atividade microbiana. As amostras foram secas ao ar e tamizadas em peneiras de 2 mm para obtenção de terra fina seca ao ar (TFSA), sendo, posteriormente, analisadas quanto ao pH em água (relação solo:água 1:2,5) e aos cátions trocáveis (Ca2+, Mg2+, K+), de acordo com os métodos descritos pela Embrapa (1997), fósforo disponível, carbono orgânico total e nitrogênio total, conforme descrito por Defelipo e Ribeiro (1981).
Os valores dessas determinações foram posteriormente relacionados com o CBM, o NBM e o PBM através de tratamento estatístico, por meio de análise de correlações: correlação de Pearson (utilizando-se o Sistema de Análises Estatísticas e Genéticas - SAEG) e análise de trilha (utilizando-se o programa Análise de Modelos Biométricos Aplicados à Genética Quantitativa e Estatística Experimental - GENES), ambos da Universidade Federal de Viçosa (UFV).
Nas análises de correlações e de trilha, os sítios da borda e do centro do platô e as amostragens do solo, da matéria orgânica leve e da manta orgânica, foram considerados isoladamente de modo a realizarem-se seis análises estatísticas independentes, em virtude de os resultados verificados por Ruivo (1991) terem mostrado que essas áreas possuem diferenças no que se refere às correlações entre as características da vegetação e as do solo. Portanto, é provável que apresentem o mesmo comportamento em relação às biomassas microbianas do solo.
A análise de trilha (desdobramento do coeficiente de correlação) consiste no estudo dos efeitos diretos e indiretos de caracteres (características explicativas) sobre uma variável básica (característica principal ou básica), dando a importância relativa do caractere na explicação da variação da variável básica (CRUZ; REGAZZI, 1994). No presente trabalho, usaram-se as características químicas determinadas no solo, na manta orgânica e na matéria orgânica leve como caracteres explicativos das variações das biomassas microbianas do solo, nos sítios alterados. Os microrganismos podem usar a matéria orgânica e os nutrientes do solo como substrato, assim como aquela fração que se encontra parcialmente decomposta, ainda na manta e pouco incorporada na matéria orgânica leve.
As características explicativas (ano de plantio; número de leguminosas; massa de matéria seca e de matéria orgânica leve; cálcio, magnésio, potássio, fósforo e nitrogênio totais, hemicelulose, celulose e lignina, na manta orgânica e na matéria orgânica leve; pH, cálcio, magnésio e potássio trocáveis, carbono orgânico, nitrogênio total, fósforo disponível e relação C/N, no solo) foram submetidas ao teste de multicolinearidade para eliminar as características que estavam inflacionando a variância. Neste estudo, eliminaram-se das análises subseqüentes as características: ano de plantio, número de leguminosas, massa de matéria seca, massa de matéria orgânica leve e hemicelulose, por apresentarem colinearidade. A análise de trilha das características (que não apresentaram colinearidade) foi utilizada para selecionar as características explicativas que mais se correlacionaram com cada uma das características indicadoras da recuperação do solo (CBM, NBM e PBM) e, posteriormente, para avaliar o efeito direto (somente o efeito positivo) das características selecionadas que estejam favorecendo o aumento das características indicadoras e, com isso, a melhoria nas condições edáficas.
RESULTADOS E DISCUSSÃO
Na Tabela 2 são apresentados os valores dos efeitos diretos e das correlações totais das características químicas da manta orgânica, da matéria orgânica leve e do solo alterado sobre os teores de C, N e P da biomassa microbiana no substrato alterado. São aqui referidos somente os elementos que apresentaram relação de causa e efeito significativa sobre os elementos da biomassa.
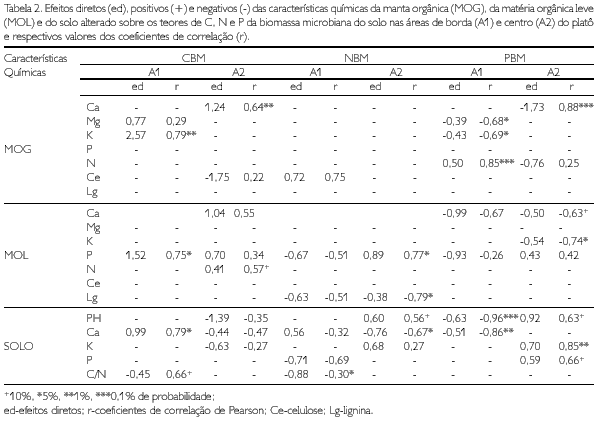
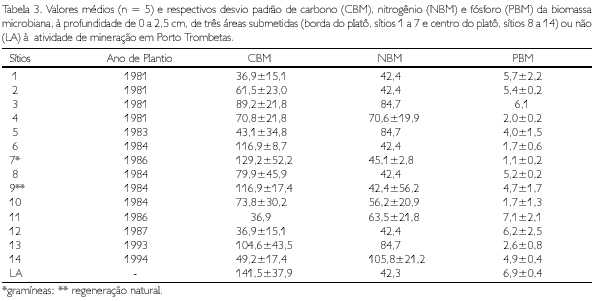
Os teores de C e P da biomassa microbiana foram menores no substrato alterado em relação ao inalterado. O nitrogênio imobilizado na biomassa microbiana do sistema inalterado, em geral, foi igual ou menor que o N imobilizado na biomassa no substrato alterado (Tabela 3). Isso pode ser de particular importância na recuperação dos sítios, visto que a biomassa microbiana representa importante compartimento de reserva de N orgânico e impede sua perda através dos processos de erosão e lixiviação.
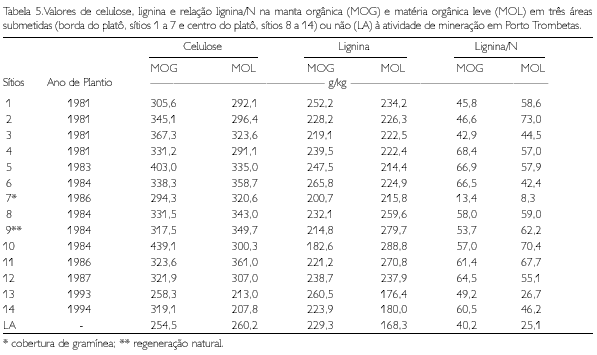
Na borda do platô, o CBM aumenta com a elevação dos teores de K+ na MOG, de P na MOL, e de Ca2+ no solo. Mesmo considerando o efeito da gramínea do sítio 7, que inflacionou os valores de K+ na MOG (já que sua folhagem foi considerada, nos cálculos, como MOG), verificou-se que os sítios 5 e 6, que mostram um dos mais altos índices de desenvolvimento da vegetação (RUIVO, 1998), apresentaram, também, altos teores de K+ na MOG (Tabela 4).
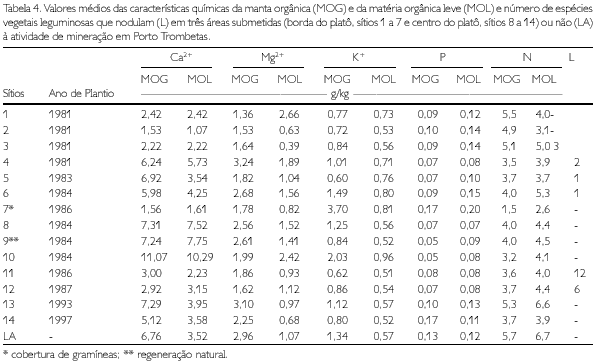
No centro do platô, o CBM eleva-se com o aumento dos teores de Ca2+ na MOG e de N na MOL. Também neste caso, os sítios que foram considerados mais próximos do LA ou em via de recuperação, assim como os sítios 10 e 13 (RUIVO, 1998) e o sítios 8, 9 e 12 apresentaram teores mais elevados de Ca2+ na MOG e N na MOL (Tabela 4).
Na borda do platô, o NBM e o PBM aumentaram, respectivamente, com a elevação nos teores de celulose (Ce) e N na MOL. Por outro lado, no centro do platô, a elevação do N na MOL e o pH do solo favoreceram o aumento do NBM. Quando se consideram o NBM e o PBM, sempre encontram-se as relações sendo confirmadas, em geral, também nos sítios 6, 9, 10 e 13 (Tabelas 4-6).
O estabelecimento, o desenvolvimento e a manutenção da população microbiana no solo dependem da existência de um substrato decomponível e do aporte constante de matéria orgânica, entre outros fatores.
Nos solos de baixa fertilidade, mas com cobertura vegetal rica em nitrogênio, como as leguminosas, menor seria a quantidade de N imobilizado pela biomassa microbiana, pois este elemento estaria em quantidade suficiente para atender à atividade metabólica dos microrganismos e ao processo de decomposição da matéria orgânica (GAMA-RODRIGUES, E.; GAMA-RODRIGUES, A. BARROS, 1997). Deve-se ressaltar que, no presente caso, os teores de N no solo alterado e na borda do platô são ligeiramente menores que no centro do platô; porém, na MOG e na MOL, os teores deste nutriente, nas duas áreas, equivalem-se.
A relação lignina/N, conforme referenciada por diversos autores (THENG; TATE; SOLLINS, 1989; ANDERSON; FLANAGAN, 1994; FRANCO; CAMPELLO, 1997), tem efeito na decomposição de resíduos vegetais. Esta relação, em geral, foi menor no material sobre o latossolo do que no solo alterado. Entre os solos alterados, a relação foi ligeiramente menor nos sítios da borda do platô (Tabela 4). Baixos valores de relação lignina/N indicam alta susceptibilidade do material à mineralização. Franco e Campello (1997) verificaram, em área degradada por mineração na Amazônia, que o teor de lignina (Lg) foi semelhante nas mantas orgânicas sob plantios de leguminosas e de Eucalyptus pellita; porém, o teor de N foi maior sob as leguminosas, resultando numa relação lignina/N menor no plantio de leguminosas do que no de eucalipto, com conseqüente maior taxa de decomposição sob as leguminosas. Assim, parece haver maior mineralização dos resíduos vegetais no solo sob vegetação primária e nas gramíneas. Na borda do platô, apesar da menor relação Lg/N, a mineralização é aparentemente mais lenta, pois não se percebe um efeito nas características químicas do solo, como esperado (Tabela 5). Em geral, os valores das propriedades físicas mostram que o solo alterado está compactado, em especial na camada de 5 a 10 cm. A compactação pela ação das máquinas (tratores e caminhões) durante a reabilitação topográfica do platô é mais pronunciada nos sítios da borda do platô (sítios 1, 2, 3, 4 e 5), fato evidenciado pelo aumento da densidade do solo e da microporosidade em relação ao LA (RUIVO, 1991).
A existência de diferenças químicas e biológicas no substrato alterado, tanto entre quanto dentro das áreas de borda e centro do platô, pode ser, em parte, devido à mistura dos horizontes do solo original durante a reabilitação da área, resultando em um substrato heterogêneo. Essas diferenças refletem-se nas necessidades ou nos requerimentos diferenciados para aumentar os elementos contidos nas células microbianas.
Uma das atividades mais importantes dos microrganismos do solo é a decomposição da matéria orgânica, com liberação de nutrientes para a solução do solo e sua absorção pelas plantas. Na borda do platô, situam-se os sítios com o substrato mineral mais pobre quimicamente, mais compactado e com maior número de espécies leguminosas. Pelo fato de nesta área também encontrarem-se os mais baixos teores de NBM, supõe-se que deve estar havendo menor quantidade de N imobilizado na biomassa microbiana.
A alteração no substrato, além da compactação, resultou também na alteração da agregação do solo, diminuindo o tamanho e a estabilidade dos agregados em água (RUIVO, 1991). Essa ação também se refletiu na diminuição da biomassa e da atividade microbiana indicados pelos teores de carbono evoluído maiores no solo inalterado (RUIVO et al., 1998). Os teores de C e P da biomassa microbiana, na profundidade de 0 a 2,5 cm, também foram mais elevados no LA, em relação ao solo alterado (Tabela 3). A maior atividade biológica no Latossolo também pôde ser verificada em lâminas delgadas. O estudo micromorfológico registra, na profundidade de 0 a 10 cm, uma intensa atividade biológica no latossolo (RUIVO, 1998) e o favorecimento da recuperação microestrutural dos solos alterados pelo retorno da matéria orgânica ao solo, seja pelo espalhamento do topsoil do LA original, seja pelo restabelecimento da cobertura vegetal que se estabeleceu com a recolonização da microbiota do solo (RUIVO et al., 2002).
A diminuição da aeração, a disponibilidade de água e a desagregação do solo, causadas pelas máquinas durante a reabilitação e constatados por Ruivo (1988), foram mais intensas na borda do platô e devem ter contribuído para diminuir a atividade biológica nesta área. Relação entre o decréscimo da atividade microbiana e aumento da compactação da camada superficial do solo foi também encontrada por Campista et al. (1996) em trilha de trator durante a remoção da cobertura vegetal, na Amazônia central.
Em solo sob pastagem, na Amazônia, Luizão, R. e Luizão, F. (1991) verificaram que, embora a biomassa microbiana fosse equivalente à da floresta, a crescente compactação do solo sob pastagem, causada pelo pastoreio, afetou a intensidade e a qualidade das atividades da microflora e da fauna, devido à diminuição da aeração e da oxigenação. A compactação desestrutura o solo, diminuindo a macroporosidade e, conseqüentemente, a fonte de água biologicamente disponível. Por outro lado, o desenvolvimento de cobertura vegetal melhora sensivelmente a estrutura física dos solos compactados, contribuindo, assim, para a recuperação da movimentação da água e dos elementos minerais no solo (CHAUVEL; GRIMELDI; TESSIER, 1990).
A conservação e a adição de MO é importante para a melhoria das características químicas (adição de C, N e P e outros nutrientes) e físicas (agregação e retenção de água) do solo e tem papel preponderante como substrato para os microrganismos. Em termos de disponibilidade de nutrientes para as plantas, os resíduos da manta e das raízes, os produtos de sua decomposição parcial e a biomassa microbiana do solo representam os componentes mais ativos da matéria orgânica e exercem papel importante na ciclagem de nutrientes.
Como o solo original é ácido e pobre em nutrientes, é evidente que o mesmo necessita de calagem e de fertilização N, P, Ke, provavelmente, de micronutrientes (que, de uma forma ou de outra, foram perdidos do solo durante as várias fases, desde a sua retirada até o seu retorno à cava) para acelerar a recuperação do solo alterado. Para isso, o manejo adequado do solo é essencial. Ele inicia-se pela retirada da cobertura vegetal e passa pelo armazenamento adequado dos horizontes do solo, principalmente do horizonte A, protegendo-o da erosão e, conseqüentemente, das perdas de MO, nutrientes e propágulos vegetais e animais. Esta proteção pode ser feita através de uma cobertura com os resíduos de parte da vegetação, o qual poderia depois ser aproveitado no recobrimento da superfície, após a reabilitação topográfica. Como abordado anteriormente, a composição química da manta orgânica e a da matéria orgânica leve relacionaram-se favoravelmente com as biomassas microbianas e, conseqüentemente, constituíram aspectos favoráveis à recuperação dos sítios alterados.
Se, com relação às características químicas, o solo alterado está mais próximo do LA, em se tratando de características biológicas o solo alterado ainda está em desequilíbrio. Embora evoluindo para a condição de equilíbrio, ele ainda está distante de ser considerado recuperado. Por isso, a biomassa microbiana apresenta-se como um indicador sensível a pequenas mudanças no ambiente e capaz de indicar o estágio de recuperação ou degradação do solo.
CONCLUSÕES
O aumento dos teores de Ca2+, K+, P, N, celulose e lignina, na manta orgânica e na matéria orgânica leve, e o de pH e dos teores de K+ e P, no solo, favoreceram, em parte, o aumento do C, N e P da biomassa microbiana do solo.
Os teores de C e P da biomassa microbiana do solo apresentaram-se adequados para separar os sítios mais degradados daqueles menos degradados, pois foram mais sensíveis às pequenas diferenças edáficas nos sítios alterados.
REFERÊNCIAS
ANDERSON, J. M.; FLANAGAN, P. W. 1994. Biological processes regulating organic matter dynamics in tropical soils. In: DORAN, J. W. et al. (Ed.). Defining soil quality for sustainable environmental. Madison. [S.l.]: SSCA, ASA. p. 97-123. (SSCA - Special Publication Number, 35).
BROOKS. P. C. et al. 1985. Chloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biology and Biochemistry, Oxford, v. 17, n. 6, p. 837-842.
BROOKS, P. C.; POWLSON, D. S.; JENKINSON, D. S. 1982. Measurement of microbial biomass phosphorus in soil. Soil Biology and Biochemistry, Oxford, v. 14, p. 319-329.
CAMPISTA, D. et al. 1996. Modificações a curto prazo no carbono, nitrogênio e biomassa microbiana no solo após o corte seletivo da floresta na Amazônia Central. In: REUNIÃO BRASILEIRA DE FERTILIDADE DO SOLO E NUTRIÇÃO DE PLANTAS, Manaus, 1996. Resumos Expandidos... Manaus: Universidade do Amazonas. p. 598-599.
CHAUVEL, A.; GRIMALDI, M.; TESSIER, D. 1990. Changes in pore space distribution following deforestation and revegetation: an example from Central Amazonian Basin, Brazil. Forest Ecology and Management, Cambridge, v. 38, p. 1-14.
CHRISTENSEN, B. T. 1992. Physical fractionation of soil and organic matter in primary particle size and density separates. In: STEWART, B. A. (Ed.). Advances in soil science. New YorK: SpringerVerlag. p. 1-90.
CRUZ, C. D.; REGAZZI, A. J. 1994. Modelos biométricos aplicados ao melhoramento genético. Viçosa: UFV, Imprensa Universitária. 390 p.
DEFELIPO, B. V., RIBEIRO, A. C. 1981. Análise química do solo (metodologia). Viçosa: UFV. 17 p. (Boletim de Extenção, 29).
DUDA, G. P. et al. 1999. Avaliação da dinâmica de matéria orgânica do solo para caracterização de áreas degradadas. Revista Brasileira de Ciência do Solo, Viçosa, v. 23, p. 723-728.
EMBRAPA; SNLCS. 1997. Manual de métodos de análises de solo. Rio de Janeiro: SNLCS.
FEIGL, B. J. et al. 1995. Soil microbial biomass in Amazonian soils: evaluation of methods and estimates of pool sizes. Soil Biology, Biochemistry, Oxford, v. 27, n. 11, p. 1467-1472.
FRANCO, A. A.; CAMPELLO, F. C. 1997. Importância da qualidade da serapilheira na sucessão vegetal em áreas de recuperação na Amazônia. In: ENCONTRO BRASILEIRO SOBRE SUBSTÂNCIAS HÜMICAS, 2., São Carlos, 1997. Anais..., São Carlos: EMBRAPA-CNPPDIA. p.53-59.
GAMA RODRIGUES, E. F.; GAMA RODRIGUES, A. C.; BARROS, N. F. 1997. Biomassa microbiana de carbono e nitrogênio de solos sob diferentes coberturas florestais. Revista Brasileira de Ciência do Solo, Viçosa, v. 21, n. 3, p. 361-365.
KÖGEL-KNABNER, I.; ZIEGLER, F. 1993. Carbon distribution in different compartments of forest soils. Geoderma, Amesterdam, v. 56, p. 241-247.
LUIZÃO, R. C.; LUIZÃO, F. J. 1991. Liteira e Biomassa Microbiana no Solo no Ciclo da Matéria Orgânica e Nutrientes em Terra Firme na Amazônia Central. In: VAL, A. L.; FIGLIUOLO, R.; FELDBERG, E. (Ed.). Bases científicas para estratégias de preservação e desenvolvimento da Amazônia, fatos e perspectivas. Manaus: INPA. v. 1. p. 65-75.
OLIVEIRA, E. P. 1997. Monitoramento da mesofauna do solo para avaliação de áreas recuperadas com árvores nativas na Mineração Rio do Norte. In: SIMPÓSIO NACIONAL DE RECUPERAÇÃO DE ÁREAS DEGRADADAS, 3., 1997, Ouro Preto. Anais... Ouro preto: SOBRADE/UFV. p. 215-222.
RUIVO, M. L. P. 1991. Caracterização das condições edáficas do Latossolo Amarelo textura muito argilosa após mineração de bauxita em Porto Trombetas-Oriximiná-Pará. 199 f. Dissertação (Mestrado em Solos e Nutrição de Plantas) - Faculdade de Ciências Agrárias do Pará, Belém.
RUIVO, M. L. P.; SCHAEFER, C. E. R. 1998. Formas e distribuição de carbono em diferentes classes de agregados em sítios recuperados após exploração de bauxita na Amazônia. In: REUNIÃO BRASILEIRA DE MANEJO E CONSERVAÇÃO DO SOLO, 12., 1998. Anais... Fortaleza: [s.n.]. p. 392-393.
RUIVO, M. L. P. 1998. Vegetação e características do solo como indicadores de reabilitação de áreas mineradas na Amazônia Oriental. 101 f. Tese (Doutorado em Solos e Nutrição de Plantas) -. Universidade Federal de Viçosa, Viçosa.
RUIVO, M. L. P. et al. 2002. Caracterização micropedológica de solos reabilitados após mineração de bauxita em Porto Trombetas (PA). Revista Brasileira de Ciência do Solo, Campinas, v. 26, n. 3, p. 759-767.
SEGAL, W.; MANCINELLI, R. L. 1987. Extent of regeneration of the microbial community in reclaimed spent oil shale land. Journal of Environmental Quality, Madison, v. 16, p. 44-47.
SOEST, P. J. Van. 1963. Use of detergents in the analysis of fibrous feeds. Journal of Association Official of Analitical Chemistry, v. 5, n. 56, p. 825-829.
THENG, B. K. G., TATE, K. R., SOLLINS, P. 1989. Constituents of organic matter in temperate and tropical soils. In: COLEMAN, D. C., OADES, J. M., UEHARA, G. (Ed.) Dynamics of soil organic matter in tropical ecosystems. Hawai: Niftal Project. p. 5-32.
TRINDADE, A. V.; TÓTOLA, M. R.; DIAS JR., H. E. 1995. Atividade microbiana em área sob impacto de mineração de ferro. In: CONGRESSO BRASILEIRO DE CIÊNCIA DO SOLO, 35., 1995, Viçosa. Resumos Expandidos... Viçosa: SBCS. v. 4. p. 2376-2378.
TURCO, R. F.; KENNEDY, A. C.; JAWSON, M. D. 1994. Microbial indicators of soil quality. In: DORAN, J. W. et al. (Ed.). Defining soil quality for sustainable environment. Madison: SSCA, ASA. p. 73-90. (SSSA Special Publication Number, 35).
VANCE, E. D.; BROOKES, P. C.; JENKINSON, D. S. 1987. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry, Oxford, v. 19, n. 6, p. 703-707.
 Endereço para correspondência:
Endereço para correspondência:
Editora do Museu Paraense Emílio Goeldi
Av. Magalhães Barata, 376
São Braz – CEP 66040-170
Caixa Postal 399
Telefone/fax: 55-91-3219-3317
E-mail:boletim@museu-goeldi.br
Recebido: 03/04/2001
Aprovado: 19/11/2002
1Parte integrante da tese de Doutorado do primeiro autor apresentada ao curso de Pós-Graduação em Solos e Nutrição de Plantas da Universidade Federal de Viçosa. Apoio Financeiro Companhia de Mineração Rio do Norte.
