










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Curriculum ScienTI
Curriculum ScienTI Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.1 n.3 Belém dez. 2006
Variação anual da produção de serapilheira em bosques de mangue no Furo Grande, Bragança, Pará
Annual variation of litterfall yield in the channel Furo Grande mangrove stands in Bragança, Pará
Adriana do Socorro da Costa Gonçalves; Marcus Emanuel Barroncas Fernandes; Muzenilha Lira Carvalho
Universidade Federal do Pará. Campus de Bragança. Instituto de Estudos Costeiros. Laboratório de Ecologia de Manguezal. Bragança, Pará, Brasil (scgonçalves.drica@bol.com.br) (mebf@ufpa.br) (muzenilha@yahoo.com.br)
RESUMO
Este estudo foi realizado no Furo Grande, Bragança, Pará, local caracterizado por três tipos de bosque de mangue: Rhizophora mangle, Avicenniagerminanse um terceiro que constitui o misto dessas duas espécies. A área de estudo apresentou uma produção de serapilheira de 7,47 t.ha-1.ano-1 (ano 1) e 7,11 t.ha-1.ano-1 (ano 2). Embora a variação nos valores mensais não tenha sido estatisticamente significativa, esta indicou uma alta produção de serapilheira nos meses do período seco (agosto a novembro de 2000) e final do período chuvoso (maio a julho de 2001). As folhas representaram, para os dois anos de estudo, em média 74% da produção total, sendo o componente com maior contribuição para a totalidade da produção em todos os sítios e durante os dois anos de trabalho, com valores que variaram de 65,5% (Sítio no3 - ano 1) a 78,4% (Sítio no1 - ano 2). Houve correlação negativa entre a produção de serapilheira e a precipitação, indicando uma estratégia de economia energética através da abscisão de folhas no período de seca. A produção de serapilheira apresentou um padrão sazonal regular, cuja tendência é se repetir nos anos subseqüentes.
Palavras-chave: Manguezal. Serapilheira. Variação sazonal. Bragança. Pará.
ABSTRACT
This study was carried out at the Furo Grande, Bragança-Pará, which is characterized by three types of mangrove stands: one dominated by Rhizophora mangle, another by Avicennia germinans, and a third one being a mixed of both species. The study site presented a litter production of 7.47 t.ha-1.year-1 (year 1) and 7.11 t.ha-1.year-1 (year 2). Although the monthly variation was not statistically significant, this variation indicated a high litter production during the dry season (August to November/2000) and the end of wet season (May to July/2001). Leaves presented an average of 74% of the total production for both years, and it was the component which presented the highest contribution to the total production in all sites during the two years of study, with values that ranged from 65.5% (Site no3 - year 1) to 78.4% (Site no1 - year 2). There was a negative correlation between the litterfall production and the precipitation, indicating a strategy of energetic economy by leaf abscission in the dry season. The litterfall production presented a regular seasonal pattern, which tendency is expected to be repeated in the subsequent years.
Keywords: Mangal. Litterfall. Seasonal variation. Bragança. Pará.
INTRODUÇÃO
Os manguezais são normalmente caracterizados pela alta produtividade primária, principalmente em locais de baixa latitude (SAENGER; SNEDAKER, 1993).
De acordo com Silva (1984), os padrões de ciclagem de nutrientes em ecossistemas florestais podem ser observados mediante uso de parâmetros de ordem temporal e estudos da quantificação dos nutrientes, os quais integram os componentes envolvidos na serapilheira, um dos componentes de maior importância no ciclo de nutrientes, sendo formada por detritos vegetais decíduos e material animal.
Para Hegazy (1998), a serapilheira tem uma importante função na cadeia alimentar, visto que permite que os ecossistemas costeiros mantenham uma atividade biológica intensa, acompanhada pela sua decomposição. Os movimentos da maré são relevantes na dispersão da serapilheira, pois viabilizam a exportação de detritos, estabelecendo uma importante função da serapilheira, que é suprir energia para as áreas estuarinas através do aporte de matéria orgânica nessas áreas (FERNANDES, 1997).
Alguns fatores bióticos parecem atuar localmente e podem estar envolvidos no processo de produtividade dos manguezais. Esses fatores são, principalmente, a salinidade (CINTRÓN et al.,1978; BALL, 1988; LIN; STERNBERG, 1992), a precipitação, a temperatura, a radiação solar, a velocidade dos ventos (DUKE; BUNT; WILLIAMS, 1984; DUKE, 1990; SLIM et al., 1996; FERNANDES, 1999; MEHLIG, 2001), as condições do solo, a limitação de nutrientes (FELLER, 1995; PEZESHKI; DELAUNE; MEEDER, 1997) e o acúmulo de gás sulfídrico ou outras substâncias tóxicas (MCKEE, 1993).
Na maioria das pesquisas sobre produtividade primária, a produção de serapilheira tem sido medida por um curto período de tempo, normalmente durante um ciclo anual. Os trabalhos desta natureza não esclarecem se a variação na produção entre as diferentes áreas estudadas é conseqüência das características específicas destas áreas ou das variações interanuais nos fatores abióticos locais e regionais, mas estudos de longo prazo ajudam a determinar o quanto essas variações na produtividade dos manguezais são resultado de uma variabilidade interanual. No presente estudo, foram analisados dados de dois anos, com o objetivo de comparar as taxas de produção de serapilheira dos bosques de mangue do Furo Grande, Bragança, Pará, para verificar as variações anuais da produção total e de seus componentes, além de comparar essas taxas de produção com os índices de precipitação local.
MATERIAL E MÉTODOS
Área de estudo
A planície costeira bragantina, no nordeste do estado do Pará, apresenta cerca de 40 km de linha costeira, estendendo-se desde a Ponta do Maiaú até a foz do rio Caeté (SOUZA FILHO, 1995). A península bragantina, localizada na costa norte do Brasil, é cortada pela rodovia PA-458, que interliga a cidade de Bragança à vila de Ajuruteua, contendo 36 km de extensão, sendo que 20 km dessa rodovia entrecortam as áreas de manguezal (CARVALHO, 2002). A área de estudo, aqui denominada de Furo Grande, localiza-se ao lado esquerdo do Km 31 dessa rodovia, no sentido norte (Figura 1), possui um comprimento de aproximadamente 12 km e é formado por pequenos canais ao lado e uma conexão parcial com o oceano Atlântico (ACHEAMPONG, 2001). Na parte mais profunda do canal, as marés de sizígia podem atingir até 7,8 m de altura, com uma velocidade de corrente de aproximadamente 2 m.s-1, que pode levar a uma alta taxa de erosão e transporte de sedimentos (DIELE, 2000).
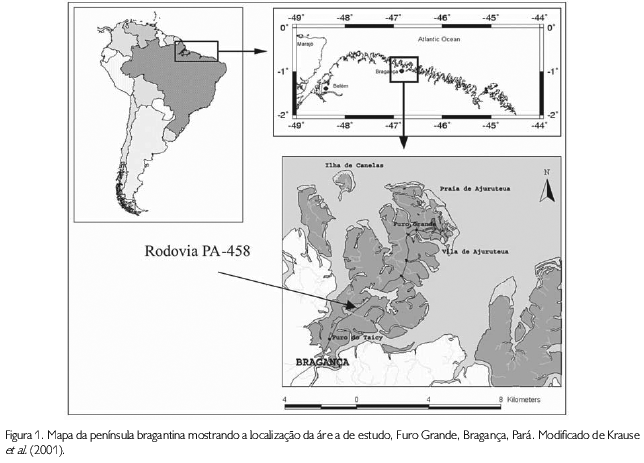
O período chuvoso na península bragantina corresponde aos meses de dezembro a junho e apresentou, durante o período de estudo, uma taxa média anual de 2.650 mm. Os valores médios mensais de precipitação (mm) utilizados neste estudo foram fornecidos pela Estação Meteorológica Automática localizada no Furo Grande (00o50'01''S - 46o38'26''W), sob a responsabilidade do Departamento de Meteorologia da Universidade Federal do Pará, Campus do Guamá.
A região na qual foram realizadas as coletas de campo caracteriza-se por apresentar manguezais bem desenvolvidos, cujas árvores atingem, aproximadamente, 20 m de altura, que pode variar de acordo com o bosque, devido aos diferentes fatores ambientais, como topografia, aporte de água e salinidade do solo (KRAUSE et al.,2001). A área de estudo escolhida foi dividida em três sítios adjacentes, sendo delimitadas com base na sua formação vegetal. O primeiro sítio (00o50 19,5 S - 46o38 14,9 W) é um bosque misto constituído de Rhizophora mangle, Avicennia germinans e Laguncularia racemosa (Sítio no1), o segundo sítio (00o50 20,7 S - 46o38 13,9 W) é um bosque puro de Avicennia germinans (Sítio no2) e o terceiro (00o50 20,9 S - 46o38 12,2 W) é um bosque dominado por Rhizophora mangle (Sítio no3).
Procedimento
Os sítios foram divididos em parcelas de 10x140 m, para sorteio e delimitação das transecções. Em cada sítio foi sorteada uma transecção de 140 m, no sentido perpendicular à rodovia e paralelo ao curso do canal e 50 m da rodovia, no intuito de eliminar efeitos de borda que poderiam ser causados pela construção da estrada (CARVALHO, 2002).
O método utilizado neste estudo foi o mesmo aplicado por Carvalho (2002). Na quantificação da serapilheira foram utilizadas cestas coletoras com moldura de madeira de 1 m2, armação de 15 cm, forradas com tela de náilon de 1 mm e suspensas permanentemente acima do nível das marés de sizígia sobre estacas de madeira. Um total de 21 cestas coletoras foram distribuídos ao longo da área de estudo, sendo sete cestas coletoras para cada sítio de trabalho, apresentando intervalos regulares de 20 m ao longo das transecções.
As coletas foram realizadas durante dois ciclos anuais completos, entre agosto de 2000 e julho de 2002. O material acumulado nas cestas foi recolhido mensalmente e estocado em sacos plásticos para separação manual em laboratório. Foram coletadas, ao longo dos dois anos, 164 subamostras nos sítios no1 e no2 e 162 subamostras no sítio no3.
No laboratório, o material coletado foi separado em folha, flor, fruto, galho, estípula e miscelânea, com os três primeiros separados por espécie. No caso do componente estípula, este só ocorre no gênero Rhizophora.Miscelânea refere-se a todo material vegetal não identificado, juntamente com fezes e outros materiais de origem animal (FERNANDES, 2003). Após a separação dos componentes, a serapilheira foi acondicionada em sacos de papel e submetida à secagem em estufa com renovação e circulação de ar a 70oC, até atingir peso constante. No final, o material foi pesado em balança de precisão para a obtenção do peso seco por componente.
Análise dos dados
A análise de variância, ANOVA - um fator, foi utilizada para comparar as taxas anuais médias de serapilheira do Furo Grande e dos sítios de trabalho. Para comparar a produção dos sítios de trabalho e dos componentes da serapilheira entre os dois anos de estudo, foi utilizada a ANOVA - dois fatores (fatorial a x b). Já os valores de produção total dos três sítios no período de dois anos foram analisados com o teste do Qui-quadrado. A variação sazonal da produção de serapilheira na área de estudo foi testada através do teste não paramétrico Kolmogorov-Smirnov (K-S) - uma amostra. A relação entre a produção de serapilheira e as taxas de precipitação foi feita através da análise de Regressão Linear Simples, com os dados de dois anos de estudo. Todas as análises foram realizadas com o auxílio do pacote estatístico Bioestat 3.0 (AYRES et al., 2003).
RESULTADOS
A produção anual de serapilheira do Furo Grande foi de 7,47 t.ha-1.ano-1 no primeiro ano e 7,11 t.ha-1.ano-1 no segundo ano. Quando analisados estatisticamente, estes valores não apresentaram uma diferença significativa (ANOVA, p>0,05).
Para a análise comparativa da produção anual de serapilheira dos três sítios de trabalho no Furo Grande foi utilizada a média geral dos dois anos de produção de cada sítio separadamente, sendo, portanto, utilizados os valores de 9,43 t.ha-1.ano-1, 6,82 t.ha-1.ano-1 e 5,63 t.ha-1ano-1 para o sítio no1, sítio no2 e sítio no3, respectivamente. De acordo com esta análise ocorreu uma diferença significativa entre os valores registrados (ANOVA, F=9,72; gl=2; p<0,001). No entanto, a comparação da produção desses três sítios nos dois anos de estudo não apresentou diferença significativa (c2= 1,92; gl=2; p>0,05). O mesmo resultado foi obtido quando realizada a análise da variação da serapilheira ao longo de um ciclo anual (K-S, p>0,05) (Figura 2).
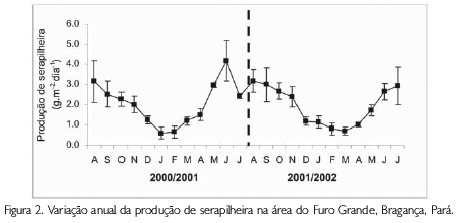
No entanto, quando a produção total de serapilheira dos três sítios foi comparada entre si, registrou-se uma diferença significativa (ANOVA, F=4,97; gl=2; p<0,01). O sítio no1, por exemplo, destacou-se por apresentar os valores mais altos nos dois anos consecutivos (9,25 t.ha-1.ano-1 e 9,60 t.ha-1ano-1), seguido pelo sítio no2 (7,68 t.ha-1.ano-1 e 6,01 t.ha-1.ano-1) e sítio no3 (5,48 t.ha-1.ano-1 e 5,78 t.ha-1.ano-1). Por outro lado, a diferença não foi significativa quando a produção dos três sítios foi comparada entre os dois anos (ANOVA, p>0,05).
Analisando os componentes separadamente, observou-se que, no primeiro ano de trabalho, o componente folha contribuiu com 72% (equivalente a 5,3 t.ha-1ano-1), flor com 8,3% (0,62 t.ha-1.ano-1), galho com 7,2% (0,54 t.ha-1.ano-1) e os outros componentes (estipula, fruto e miscelânea) com 12,5% (0,9 t.ha-1.ano-1) (Figura 3).
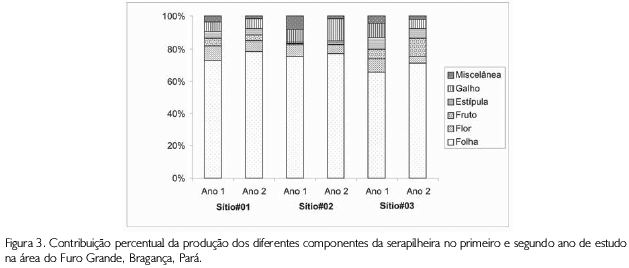
No segundo ano, folha, flor e galho também foram os componentes de maior contribuição, com 76% (5,42 t.ha-1.ano-1), 5,4% (0,39 t.ha-1.ano-1) e 8,27% (0,59 t.ha-1.ano-1), respectivamente, ao passo que fruto, estipula e miscelânea contribuíram com apenas 10,1% (0,72 t.ha-1.ano-1) (Figura 3). Quando comparados entre si, estes componentes apresentaram diferença significativa (ANOVA, F=52,56; gl=5; p<0,001); em contrapartida, a diferença de produção desses componentes entre os dois anos de estudo não foi significativa (ANOVA, p>0,05).
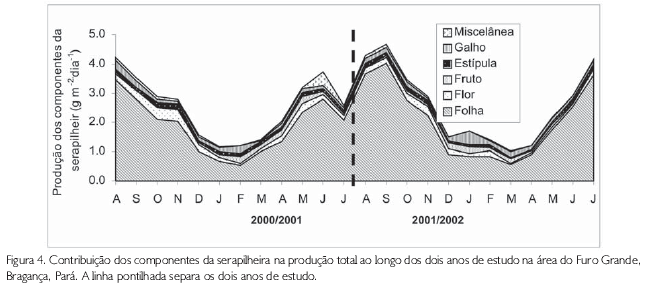
Considerando a produção entre os componentes e os sítios, pode-se observar que ocorreu uma diferença significativa para os três bosques (Misto - ANOVA, F = 63,27; gl=5; p<0,001; Avicennia - ANOVA, F=26,70; gl=5; p<0,001; e Rhizophora - ANOVA, F=42,30; gl=5; p<0,001). Para esta produção, observou-se, também, que a diferença não foi significativa quando a comparação é feita entre os anos (ANOVA, p>0,05). Folha apresentou a maior contribuição para a produção total em todos os sítios e durante os dois anos de trabalho, com valores que variaram de 65,5% (sítio no 03 - ano 1) a 78,4% (sítio no 01 - ano 2). A variação mensal não apresentou diferença significativa entre os meses do período de estudo (K-S, p>0,05) (Figura 4).
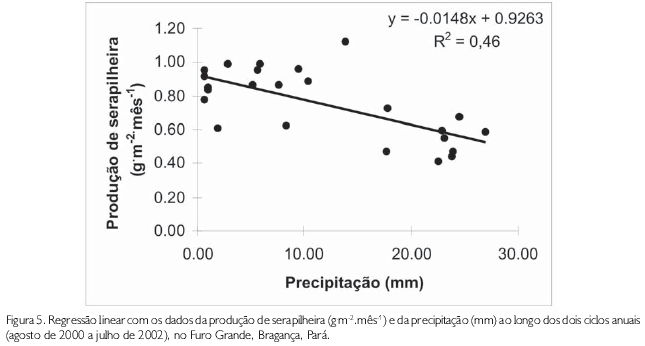
Uma análise de regressão linear foi utilizada para investigar a relação entre a produção de serapilheira e a precipitação. Foram utilizados os valores dos dois anos de estudo. O valor da correlação de Pearson (r) encontrado foi de 46%, apresentando uma tendência negativa e significativa (RL, F=19,23; gl=11; p<0,001) (Figura 5).
DISCUSSÃO
A maioria dos estudos sobre a produção de serapilheira do ecossistema manguezal na Amazônia brasileira foi desenvolvida nos bosques de mangue da península bragantina. Isto permite que os resultados aqui apresentados sejam comparados com os já existentes para a mesma região. Por exemplo, o estudo anteriormente realizado no Furo Grande por Carvalho (2002) também registrou valor aproximado ao encontrado no presente trabalho (7,8 t.ha-1.ano-1); valores superiores foram estimados por Mehlig (2001), nas áreas de manguezal do Acarajó e Furo do Meio (12,8 e 13,1 t.ha-1.ano-1, respectivamente); além disso, estimativas de diferentes bosques de mangue (Rhizophora, Avicennia e Laguncularia) registraram valores desde 2,3 a 16,5 t.ha-1.ano-1 na ilha de Maracá, estado do Amapá (FERNANDES, 2003), cujos valores, segundo este autor, são comparáveis ou mesmo maior do que aqueles dos mangues de outros países.
De acordo com os resultados deste estudo para os bosques de Rhizophora e Avicennia, a produção de serapilheira foi menor do que aquela dos bosques dominados pelos mesmos gêneros botânicos da ilha de Maracá (14,1 t.ha-1.ano-1 e 7,3 t.ha-1.ano-1) (FERNANDES, 2003). Por outro lado, os valores aqui apresentados para os bosques misto e de Avicennia são maiores do que aqueles estudados por Farias (2003) para os bosques misto e anão na península bragantina, cujos valores foram de 4,93 t.ha-1.ano-1 e 1,89 t.ha-1.ano-1, respectivamente. Diferenças na produtividade desses bosques está, provavelmente, relacionada ao padrão estrutural que eles apresentam, principalmente o bosque anão, que se caracteriza por apresentar estrutura arbustiva, em conseqüência da alta salinidade do solo intensificada durante a estação seca (MEDINA et al.,2001). Os bosques do Furo Grande, segundo Seixas (2003), apresentam estrutura mais desenvolvida, com altura média de 10,9 m (bosque de Avicennia) e 10,2 m (bosque Misto), o que proporciona a produção de um maior volume de serapilheira para o ecossistema.
A variação dos valores mensais da produção de serapilheira no Furo Grande não foi estatisticamente significativa, mas esta variação indicou uma alta produção de serapilheira nos meses do período seco (agosto a novembro de 2000) e final do período chuvoso (maio a julho de 2001), concordando com os dados apresentados por Fernandes (1997), Mehlig (2001) e Carvalho (2002).
Os resultados obtidos para averiguar a contribuição dos componentes da serapilheira concordam com aqueles obtidos por Fernandes (1997), Mehlig (2001) e Carvalho (2002). Os valores de produção de serapilheira dos bosques do presente estudo foram diretamente influenciados pelas taxas de queda do componente folha, considerada como o mais importante componente da serapilheira, uma vez que atingem altas porcentagens da produção total (POOL; LUGO, SNEDAKER, 1975; SASEKUMAR; LOI, 1983; FERNANDES, 1997; MEHLIG, 2001; BATISTA, 2003; FARIAS, 2003).
A queda de serapilheira na área do Furo Grande foi variável ao longo do período deste estudo. Nos dois anos, a produção parece aumentar por volta de abril, entretanto, foram observadas variações com relação aos picos de produção: junho, por exemplo, foi o mês mais produtivo no primeiro ano, enquanto no segundo o pico ocorreu em agosto. De fato, existe uma correlação entre a produção de serapilheira na área de estudo e o período seco, muito embora o valor estatístico seja baixo. Contudo, esses resultados apontam para um padrão geral observado nas florestas tropicais, onde o pico da produção de serapilheira é registrado no período seco, o que pode estar relacionado com o balanço energético das plantas, haja vista a abscisão de folhas, maior componente da serapilheira, representar uma economia de energia nos períodos de maior estresse hídrico (AMARASINGHE; BALASUBRAMANIAM, 1992).
De acordo com Saenger e Snedaker (1993), a produção de serapilheira é maior próximo à linha do equador. No entanto, é importante ressaltar que a produção de serapilheira é variável quando se considera uma escala espacial pequena, sugerindo que as espécies de mangue reagem às condições locais, como parece ser o caso dos bosques de mangue do presente estudo. Por fim, é relevante enfatizar que os resultados aqui apresentados também apontam para um padrão sazonal regular quando mais de um ciclo anual é avaliado, ou seja, o padrão tende a ser repetido nos anos subseqüentes.
REFERÊNCIAS
ACHEAMPONG, E. 2001. Distribution of macrozoobenthos abundance and biomass in intertidal soft sediments of Northeast Brazil. Dissertation (MSc.) - University of Bremen, Bremen.
AMARASINGHE, M. D.; BALASUBRAMANIAM. 1992. Net primary productivity of two mangrove forest stands on the northwestern coast of Sri Lanka. Hydrobiologia, v. 247, p. 37-47.
AYRES, M. et al. 2003. BioEstat 3.0: aplicações estatísticas nas áreas das ciências biológicas e médicas. Belém: Sociedade Civil Mamirauá.
BALL, M. 1988. Ecophysiology of mangroves. Trees, v. 2, p. 129-142.
BATISTA, B. S. 2003. Produção de um bosque de Avicennia sob um gradiente de: inundação e salinidade na península Bragantina. Monografia (Trabalho de Conclusão de Curso) - Universidade Federal do Pará, Campus de Bragança, Bragança-Pará.
CARVALHO, M. L. 2002. Aspectos da produtividade primária dos bosques de mangue do Furo Grande, Bragança-Pará. Dissertação (Mestrado) - Universidade Federal do Pará, Belém-Pará.
CINTRÓN, G. et al. 1978. Mangroves of arid environments in Puerto Rico and adjacent islands. Biotropica, v. 10, p. 110-121.
DIELE, K. 2000. Life history and population structure of the exploited mangrove crab Ucides cordatus (Linnaeus, 1763) (Decapoda: Brachyura) in the Caeté estuary, North Brazil. [S.l.:s.n.]. 103 p. (ZMT Contribution, 9).
DUKE, N. C. 1990. Phenological trends with latitude in the mangrove tree Avicennia marina. Journal of Ecology, v. 78, p. 113-133.
DUKE, N. C.; BUNT, J. S.; WILLIAMS, W. T. 1984. Observations on the floral and vegetative phenologies of north-eastern Australian mangroves. Australian Journal Botany, v. 32, p. 87-99.
FARIAS, A. S. C. 2003. Comparação da produção de serapilheira de dois bosques de mangue com diferentes padrões estruturais na península bragantina, Bragança-Pará. Monografia (Trabalho de Conclusão de Curso) - Universidade Federal do Pará, Campus de Bragança, Bragança-Pará.
FELLER, I. C. 1995. Effects of nutrient enrichment on growth and herbivory of dwarf red mangrove (Rhizophora mangle). Ecological Monographs, v. 65, p. 477-505.
FERNANDES, M. E. B. 1997. The ecology and productivity of mangroves in the Amazon region, Brazil. Tese (Doutorado) - University of York, England.
FERNANDES, M. E. B. 1999. Phenological patterns of Rhizophora L., Avicennia L. and Laguncularia Gaertn. f. in Amazonian mangrove swamps. Hydrobiologia, v. 413, p. 53-62.
FERNANDES, M. E. B. 2003. Produção primária: Serapilheira. In: FERNANDES, M. E. B. (Ed.). Os manguezais da costa norte brasileira. São Luís-MA: Fundação Rio Bacanga. p. 61-78.
HEGAZY, A. K. 1998. Perspectives on survival, phenology, litter fall and decomposition, and caloric content of Avicennia marina in the Arabian Gulf region. Journal of Arid Environments, v. 40, p. 417-429.
KRAUSE, G. et al. 2001. Spatial Patterns of Mangrove Ecosystems: The Bragantinian Mangroves of Norther Brazil. Ecotropica, v. 7, p. 93-170.
LIN, G.; STERNBERG, L. S. L. 1992. Differences in morphology, carbon isotope ratios, and Photosynthesis between scrub and fringe mangroves in Florida, USA. Aquatic Botany, v. 42, p. 303-313.
MCKEE, K. L. 1993. Soil physicochemical patterns and mangrove species distribution - reciprocal effects? Journal of Ecology, p. 81, p. 477-487.
MEDINA, E. et al. 2001. Mangal communities of the "Salgado Paraense": Ecological heterogeneity along the Bragança península assessed through soil and leaf analyses. Amazoniana, v. 16, p. 397-416.
MEHLIG, U. 2001. Aspects of tree primary production in an equatorial mangrove forest in Brazil. Bremen: Center for Tropical Marine Ecology (ZMT). 155 p. (ZMT contribution, 14).
PEZESHKI, S. R.; DELAUNE, R. D.; MEEDER, J. F. 1997. Carbon assimilation and biomass partitioning in Avicennia germinans and Rhizophora mangle seedlings in response to soil redox conditions. Environmental and Experimental Botany, v. 37, p. 161-171.
POOL, D. J.; LUGO, A. E.; SNEDAKER, S. C. 1975. Litter production in mangrove forests of southern Florida and Puerto Rico. In: WALSH, G. E.; SNEDAKER, S. C.; TEAS, H. J. (Ed.). Proc. Int. Symp. Biol. and Management of Mangroves, Univ. Florida: IFAS. p. 213-299.
SAENGER, P.; SNEDAKER, S. C. 1993. Pantropical trends in mangrove above-ground biomass and annual litterfall. Ecologia, v. 96, p. 293-299.
SASEKUMAR, A.; LOI, J. J. 1983. Litter production in three mangrove forest zones in the Malay Peninsula. Aquatic Botany, v. 17, p. 283-290.
SEIXAS, J. A. S. 2003. Estudo comparativo de vegetação dos bosques de mangue do Furo Grande Bragança-Pará. Monografia (Trabalho de Conclusão de Curso) - Universidade Federal do Pará, Campus de Bragança.
SILVA, M. F. F. 1984. Produção anual de serapilheira e seu conteúdo mineralógico em mata tropical de terra firme, e várzea e igapó. Boletim Museu Paraense Emílio Goeldi. série Botânica, v. 56, p. 1-13.
SLIM, F. J. et al. 1996. Biomass and litterfall of C. tagal and R. mucronata in the mangrove forest of Gazi Bay, Kenya. Mar. Freshwater Res. v. 47, p. 999-1007.
SOUZA FILHO, P. W. M. 1995. Influência das variações do nível do mar na morfoestratigrafia da planície costeira bragantina (NE do PA) durante o Holoceno. Dissertação (Mestrado) - Universidade Federal do Pará, Belém-Pará.
 Endereço para correspondência:
Endereço para correspondência:
Editora do Museu Paraense Emílio Goeldi
Av. Magalhães Barata, 376
São Braz – CEP 66040-170
Caixa Postal 399
Telefone/fax: 55-91-3219-3317
E-mail: boletim@museu-goeldi.br
Recebido: 10/03/2005
Aprovado: 27/03/2006
