










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.2 n.1 Belém abr. 2007
Osteologia craniana e considerações sistemáticas em Coccyzinae (Cuculidae)
Cranial osteology and systematic considerations in Coccyzinae (Cuculidae)
Sérgio Roberto PossoI; Reginaldo José DonatelliII
IUniversidade Federal do Mato Grosso do Sul. CPTL/DCN. Três Lagoas, Mato Grosso do Sul, Brasil srposso@hotmail.com
IIUniversidade Estadual Paulista. Campus de Bauru. Faculdade de Ciências. Departamento de Ciências Biológicas. Laboratório de Vertebrados. Bauru, São Paulo, Brasil rjdonat@yahoo.com.br
RESUMO
Pouco se conhece sobre a osteología da subfamilia Coccyzinae e não há consenso sobre sua monoíilia e posicionamento dentro dos Cuculidae. Sendo assim, descreveu-se detalhada e comparativamente a osteología de 86 espécimes pertencentes a 11 espécies, abrangendo todos os quatro gêneros de Coccyzinae. Os crânios cos Coccyzinae são morfologicamente homogêneos nas regiões frontal, parietal, occipital, paraesfenóide e ossos lacrimal e quadrado; mas outras estruturas e regiões, como a maxila superior, região esquamosal, palato, ectetmóide e mandíbula, apresentam variações morfológicas, principalmente em nível genérico. Estas variações fornecem, a priori, algumas relações de similandade entre ostáxons de Coccyzinae, principalmente entre Coccyzuse Hyetornis, As espécies congêneres pouco apresentam modificações quando comparadas entre si, à exceção de Playa minuta, claramente distinta das demais Piaya em 11 características, o que fornece indícios para uma nova investigação sobre a validade e posicionamento desta espécie dentro do gênero Piaya. Os caracteres osteológicos cranianos obtidos por meio deste estudo serão aplicados em uma futura análise filogenética para teste das hipóteses aqui levantadas e de esclarecimento das divergências taxonómicas desta controversa subfamilia.
Palavras-chave: Cuculiformes. Coccyzinae. Crânio. Osteología. Sistemática.
ABSTRACT
There s no consent whether the Coccyzinae is a monophyletic group and also about its position within Cuculidae. Furthermore, the osteology is poorly studied in this group. Thus, we described the cranial osteology of the 86 specimens belonging to 11 species in all four genera of Coccyzinae. The skulls are comparatively homogeneous in relation to the frontal, parietal, occipital, paraesphenoid regions and lacnmal and quadrate bones; however the upper jaw, squamosal region, palate and ectethmoid bones and mandible present morphologic variations mainly in generic level. These vanations supply, a priori, some similarity relationships among the Coccyzinae taxa, mainly between Coccyzus and Hyetornis. The species within each genera are very similiar to each other, the exception is Piaya minuta that shows 11 diferent characteristics from other Piaya species. This result demands further investigations whether P. minuta should be considered within Piaya. The cranial osteological characters described here will be useful for a future cladistic analysis in order to test the hypotheses raised and also to solve the problematic classification of this controversial group.
Keywords: Cuculiformes. Coccyzinae. Skull. Osteology. Systematic.
INTRODUÇÃO
A subfamília Coccyzinae atualmente é composta por quatro gêneros e 18 espécies, com nove espécies atribuídas ao gênero Coccyzus, três para Piaya, quatro para Saurothera. sendo Hyetomis drtípico (PAYNE, 1997).
As espécies pertencentes a este grupo ocorrem principalmente na região Neotropical, com exceção de Coccyzus americanus e Coccyzus erythrophtalmus, que migram entre as Améncas do Sul e Norte. Os gêneros Saurothera e Hyetornis são endêmicos de ilhas do Caribe, enquanto Piaya e Coccyzus estão amplamente distribuídos pelas Américas Central e Sul (SIBLEY; MONROE, 1990).
Não se observam características exclusivas para o grupo na literatura. Basicamente são arbóreos, constróem ninhos e ocupam diversos habitats, incluindo florestas tropicais, florestas secundárias, mangues, cerrado e campos abertos (HUGHES, 1996; PAYNE, 1997).
Feduccia (1996) afirma que Foro panarium possui características atribuídas tanto aos cucos quanto aos turacos (Musophagidae) e à cigana (Opisthocomus hoazin), além de ser o fóssil mais antigo relacionado aos Cuculiformes datado do Eoceno (57 milhões anos) na América do Norte. Payne (1997) relata que Neococcyx mccorquodalei, com características exclusivas dos cucos e provavelmente pertencente à subfamflia Coccyzinae, é datado do Oligoceno Inferior no Canadá, indicando que este grupo ocorre nesta região há, no mínimo. 36 mlhões de anos.
A monofiüa e posição filogenética desta subfamlia é objeto de controvérsia entre os sistematas. Segundo Seibel (1988) e Hughes (1996, 2000). os Coccyzinae são polrfiléticos (Coccyzus e Saurothera estão distantemente relacionados de Piaya e Hyetornis)e estão mais próximos dos Cuculidae do Velho Mundo (Cuculinae). Sibley e Ahlquist (1990) e Payne (1997) consideram-no um grupo monofilético; ainda, Sibley e Ahlquist, assim como Seibel (1988) e Hughes (1996, 2000), aproximam-nos dos Cuculidae do Velho Mundo. Entretanto, este posicionamento é preterido por Payne (1997) que, com base em dados moleculares e eletroforese de queratina das penas (BRUSH; VvTTT, 1983), inclui os Coccyzinae, juntamente com os demais cucos do Novo Mundo (Crotophaginae e Neomorphinae), em um grupo distinto dos cucos do Velho Mundo.
Outro assunto bastante controverso na literatura refere-se ao padrão evolutivo do parasitismo de ninho nos Cuculiformes, uma vez que pode ter surgido duas vezes independentemente nesta ordem (PAYNE, 1997; SICK, 1997) ou, de acordo com os cladogramas obtidos em Seibel (1988) e Hughes (1996, 2000), admite-se somente um surgimento de parasitismo nos Cuculiformes. O posicionamento filogenético dos Coccyzinae poderá auxiliar na tentativa de esclarecer tais divergências, já que as espécies de Coccyzus apresentam parasitismo de ninho facultativo.
Em relação à anatomia, Berger (1960) aponta a necessidade de maiores estudos anatômicos com os cucos em geral, particularmente em relação ao crânio e mandíbula, para que seja possível fornecer maiores subsídios para o refinamento sistemático deste controverso grupo.
Tendo apontado tais lacunas em relação à osteología e à sistemática da subfamilia Coccyzinae, objetivou-se neste estudo: descrever detalhada e comparativamente a osteología craniana das espécies pertencentes aos Coccyzinae, de acordo com a classificação de Payne (1997), visando a contribuir para o refinamento da anatomia neste grupo; e. por meio do levantamento de caracteres osteológicos cranianos, procurou-se estabelecer as possíveis relações de similaridade morfológica entre as espécies e gêneros aqui considerados, além de se obter caracteres para serem utilizados em uma futura análise cladística.
MATERIAL E MÉTODOS
Estudou-se a anatomia óssea craniana em 86 espécimes pertencentes a 11 espécies agrupadas nos quatro gêneros pertencentes à subfamilia Coccyzinae. Os espécimes analisados fazem parte das coleções do Museu Paraense Emílio Goeldi (MPEG), Belém, Brasil; Museum of Zoology (UMMZ), University of Michigan, Ann Arbor, Michigan, EUA; United States National Museum (USNM), Smithsonian Insttution, Washington, EUA; e do Museu de História Natural de Tãubaté, Brasil (MHNT).
Segue-se a relação do material estudado: Coccyzus americanus (US|NM: 554343, 501301, 289834, 501300, 17694, 553294, 557820, 19288, 614143, 17685; MHNT: 455, 608); C. erythrophtalmus (USNM 288242, 345450, 288240, 499620, 288241, 224427, 346327, 377, 6560, 224424; MHNT: 562); C. meiacoryphus (USNM: 612039, 227775; MHNT: 1410, 251); C. minor (USNM: 557692, 555759, 555758, 555756, 555000, 555001, 558085, 558083, 555535, 553323; MHNT: 547); Hyetornis pluvialis (USNM: 559183, 558866, 558867, 559182); H. rufigularis (USNM: 554608, 226541, 226540); Piaya cayana (USNM: 559338, 344101, 613974, 492313, 559337, 558792, 289792, 347111, 288783, 345878; MPEG: 1116, 1483, 1163; MHNT: 112, 395, 1140); P. melanogaster (USNM: 559339); P. minuta (USNM: 345882, 345881, 345883, 432599; MHNT: 763); Saurothera merlini (USNM: 427940, 553324, 555173, 429287, 429288, 19645; UMMZ: 1 58527); S. vetula (USNM: 501735, 292568, 555763, 501792, 501870, 292570, 554671, 554610, 555760, 292567, 501920, 501704).
As delimitações das regiões do crânio foram baseadas na descrição inicial de Piaya cayana, sendo tais regiões apontadas nas ilustrações desta espécie, bem como a maioria das estruturas ósseas. Tais delimitações serviram, ainda, de parâmetro para comparação com os demais espécimes estudados.
A osteologia craniana foi descrita e estudada comparativamente com auxílio de um estereomicroscópio Leika Wild M3B (9,6x- 60x). Para as medidas das estruturas ósseas utilizou-se um paquímetro Mitutoyo de 300 mm e as proporções relativas apresentadas são resultado de uma média de três mensurações da mesma estrutura em cada crânio.
Por haver fusão entre a maioria dos ossos do crânio e da mandíbula nas Aves, nem sempre se pode caracterizar os limites ósseos de algumas estruturas. Assim, muitas estruturas são aqui descritas através do uso de limites determinados por regiões, seguindo os trabalhos de Hõfling e Gasc (1984), Hõfling (1995) e DonateUi (1992. 1996, 1997).
São apresentadas ilustrações, acompanhadas de legendas, para facilitar as observações e comparações das estruturas descritas. Para melhor localizar as estruturas ósseas descritas nas diversas espécies de Coccyzmae, em cada espécie é indicada, ainda, a figura em que se encontra determinada estrutura. Nas vistas caudal do crânio e mandíbula é apresentada somente uma ilustração representativa para cada gênero, uma vez que não apresentam variação intragenérica.
A Tabela 1 tem porfmalidade apresentar um resumo para comparação das diversas estruturas ósseas cranianas que apresentaram morfologia heterogênea entre as espécies de Coccyzinae.
Quanto à nomenclatura (terminologia) utilizada para a descrição da osteologia. seguiu-se aquela proposta pela Nomina Anatômica Avium (BAUMEL; WITMER, 1993). Nos casos em que não foi possível utilizar esse referencial, optou-se pela utilização de letras e números a fim de evitar a criação desnecessária de nomes, seguindo Hõfling e Gasc (1984), Hõfling (1995) e Donatelli (1992, 1996, 1997).
Para evitar a criação de sinonimias em relação às estruturas anatômicas, a Nomina Anatómica Avium (BAUMEL ef a/., 1993) denomina as estruturas em latim. Todavia, tal procedimento dificulta a compreensão de estudos anatômicos descritos em língua portuguesa. Visando a minimizar este problema, a nomenclatura osteológica está traduzida para a língua portuguesa, contudo, apresenta-se o correspondente em latim uma única vez quando cada estrutura óssea é descrita no texto.
A nomenclatura e a classificação das espécies é aquela apresentada por Payne (1997), uma vez que se objetiva testar esta hipótese de classificação em uma futura análise filogenética.
RESULTADOS
Osteología
Ossa cranii
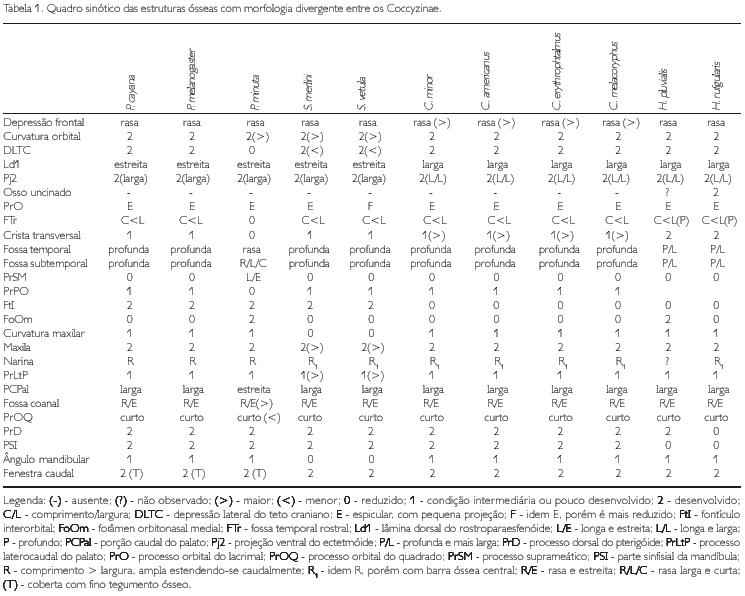
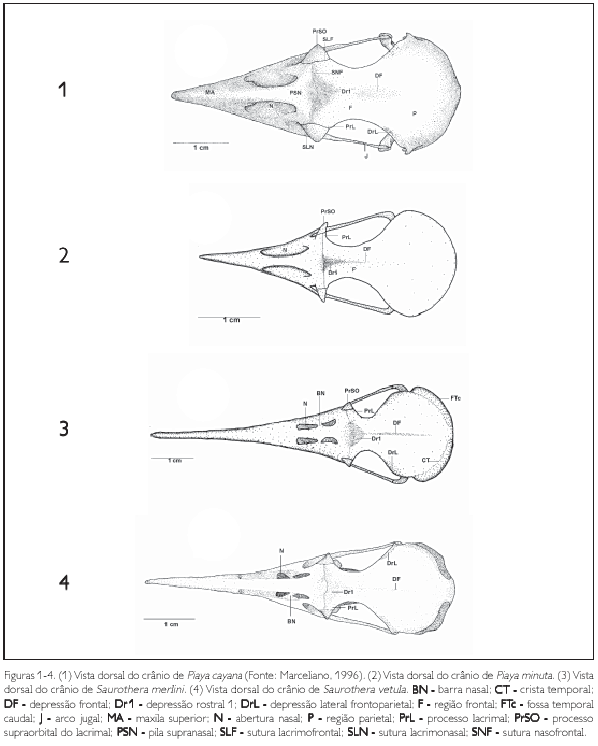
A região frontal (os frontale) (F: Figura 1) limita se rostrolateralmente com o osso lacrimal através da sutura lacrimofrontal (SLF: Figuras 1, 5 e 9); rostralmente, na zona flexoria craniofacialis, articula se com a maxila superior por meio da sutura nasofrontal (5NF: Figuras 1, 5 e 9), sendo que estas regiões se sobrepõem entre si (tendência à sinartrose), fazendo com que a sutura frontonasal seja pouco conspícua. Ventralmente, a região frontal funde se ao septo interorbital e, em sua porção caudal, funde se com a região parietal. Na zona flexoria craniofacialis, todas as espécies apresentam a face rostral da região frontal formando uma depressão rostral 1 (Drl: Figuras 1 -4, 5-8, 9 e 10). Na facies articuiaris frontonasalis (região da sutura com o osso lacrimal), observa se o processo lacnmal (proc. lacrimalis) (PrL: Figuras 1 a 10), largo lateralmente em decorrência da menor largura lateral na porção orbital da região frontal. Observa se na região frontal um conspícuo sulco mediano (depressio frontalis) (DF: Figuras 1 a 10) que se estende até a região parietal, sendo este sulco mais profundo nas espécies de Coccyzus (Figuras 5 8). Todas as espécies apresentam, na porção orbital, as extremidades laterais da região frontal curvadas (côncavas) medialmente e são mais estreitas se comparadas à extensão lateral da abóbada craniana (Calvaria). P. minuta, S. merlini e S. vetula (Figuras 2-4) apresentam esta curvatura acentuada, em que a extensão lateral da abóbada craniana possui cerca de três vezes mais a largura da expansão lateral frontal. Nas espécies pertencentes aos gêneros Coccyzus (Figuras 5-8), Hyetornis (Figuras 9 e 10) e Piaya cayana (Figura 1), esta diferença atinge cerca de 2,5 vezes a mesma medida.

A região parietal (os panetalé) (P: Figuras 1 e 11) encontra se totalmente fundida rostralmente com a região frontal; rostrolateralmente, contata se com a fossa temporal através das cristas temporais 1 e 2; caudalmente com as cnstas nucal tranversa (crista nuchalis transversa) (CNT: Figuras 11-14) e nucal sagital (crista nuchalis sagitalis) (CNS: Figuras 11 -14). Todas as espécies apresentam uma rasa depressão nas extremidades das bordas laterais da porção rostrolateral do teto craniano (DrL: Figuras 1-4, 5-8, 9 e 10 e 14), dorsorostralmente em relação ao processo pós orbital, entretanto, em P. minuta (Figura 2) esta é extremamente rasa e, em S. merlini (Figura 3) e S. vetula (Figura 4), tal depressão é pouco conspícua em relação às demais.
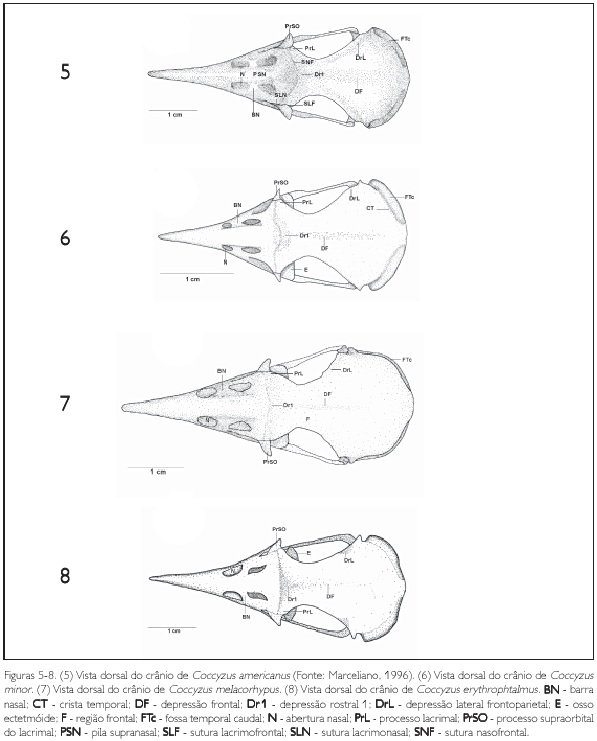
A região occipital compreende as regiões dos ossos supra occipital (os supraoccipitale), exoccipital (os exoccipitale)e basiocciprtal (os basioccipitale) fundidas. Toda esta região encontra se aprofundada no crânio, em relação às regiões panetal e esquamosal, fazendo com que a crista nucal transversa seja mais proeminente. O forâmen magno (foramen magnum) (FoMa: Figuras 11-14) apresenta se em forma circular em todas as espécies descritas e está delimitado pelas três regiões supracitadas.
A região supra occipital (SO: Figura 11) continua-se ventrolateralmente com a região exoccipital. Essa região apresenta, em sua porção medial, uma elevação óssea característica (proeminentia cerebeJaris) (PrC: Figuras 11-14) que se encontra pouco desenvolvida em todas as espécies observadas. Dorsolateral mente em relação ao foramen magno, em alguns espécimes é possível se observar o foramen da veia occipital externa (foramen venum occipitalis externae) (FoVOE: Figuras 11-13 e 33) pouco conspícuo.
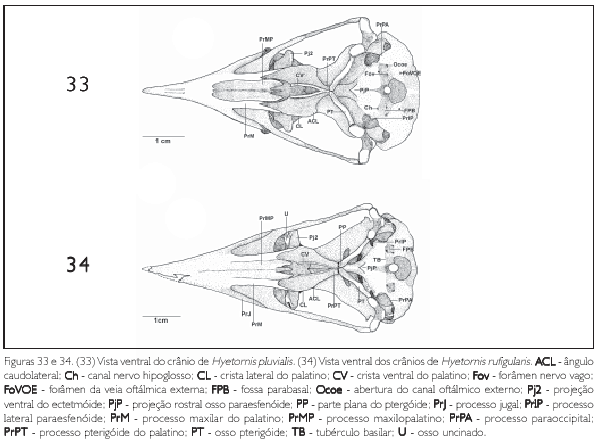
A região exoccipital (EX: Figura 11) é delimitada medioventralmente pela região basioccipital e laterorostralmente pelo meato acústico externo por meio do processo paraoccipital (proc. parocàpitalis) (PrPA: Figuras 11-14, 25-28, 29-32 e 33). Este processo projeta se ventralmente com ápice arredondado e encontra se desenvolvido em todas as espécies descritas. Na fossa parabasal (fossa parabasalis) (FPB: Figuras 25-28, 29 32, 33 e 34) da região exoccipital, lateralmente à região basiesfenóide, medialmente em relação ao meato acústico, encontram se quatro aberturas: da veia carótida (ostium canaJis carotid) (OCC: Figura 29), relativamente próxima à região basiesfenóide; do nervo oftálmico externo (ostium canaiis ophthalmia externi) (Ocoe: Figuras 25, 27 a 30 e 33), situado próximo ao processo parocciprtal, sendo a abertura com o maior calibre entre as aberturas da fossa parabasal; foramen do nervo vago (foramen nervus vagi) (Fov: Figuras 25, 30 e 33), situado mediocaudalmente em relação às aberturas supracitadas, apresentando o menor calibre dentre as descritas; e canal do nervo hipoglosso (canalis nervus hypogiosst) (Ch: Figuras 25, 27, 28, 29, 30 e 33) localizado rostromedialmente em relação às aberturas supracitadas. O desenvolvimento do calibre das aberturas destes forâmens supracitados formam um padrão característico entre as espécies estudadas, isto é, não há variações na forma e posição de tais aberturas entre as espécies.

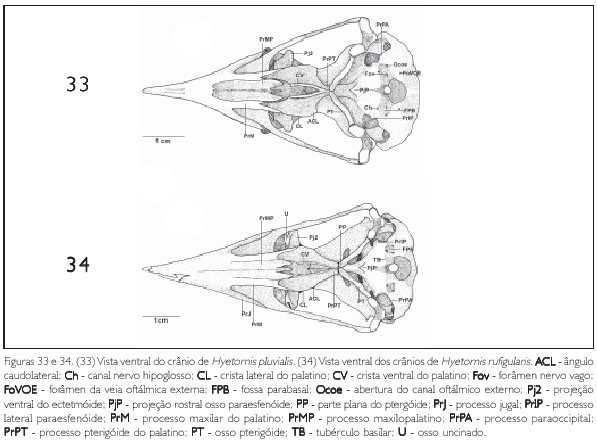
Em sua porção rostral, a região basioccipital (B: Figura 25) contata se com a região basiesfenóide através do tubérculo basilar (tuberculum basilaré) (TB: Figuras 25, 26, 30, 32 e 34) pouco proeminente e em forma de uma cnsta. Em sua porção caudal observa se o côndilo occipital (œndylus occipitalis) (COC: Figuras 11 e 25) com sua respectiva incisara mediana condyli, situada em sua porção caudal. Este côndilo. bem como sua incisura, é pouco pronunciado nas espécies descritas. Afossa subcondilar (fossa subcondilaris) (FS: Figuras 26 e 29) circunda rostroventralmente o côndilo occipital e encontra se rasa em todas as espécies observadas.
A região basiesfenóide (os basisphenoidale) (Ba: Figura 25) situa-se rostralmente à região basioccipital, lateralmente limita se com o meato acústico externo e rostralmente funde se à região paraesfenóide.
A região paraesfenóide (os parasphenoidale) (OS: Figura 25) é laminar e sua base rostral (basis rostri parasphenoidale)projeta-se rostralmente, recobrindo parte da face caudal do rostro paraesfenóide através da projeção paraesfenóide (PjP: Figuras 28, 31, 32, 33 e 34), que se apresenta pouco desenvolvida afilando se em sua extremidade rostral com ápice pontiagudo. O processo lateral paraesfenóide (proc. lateralis parasphenoidale) (PrlP: Figuras 11, 25-28 e 29, 31, 32, 33 e 34) projeta se ventralmente e encontra se reduzido e afilado ventralmente. Todas as espécies estudadas não apresentam o processo medial paraesfenóide (proc. medialis parasphenoidalis).
O rostro paraesfenóide (rostrum parasphenoidale) (RP: Figura 25) projeta se rostralmente, emite dorsalmente uma lâmina 1 delgada (Ld1: Figuras 15, 20 e 23) até os limites com o fontículo interorbital, fundindo se rostralmente à região do osso mesetmóide e septo interorbital. Nas espécies pertencentes aos gêneros Coccyzus (Figuras 19-22) e Hyetornis (Figuras 23 e 24) esta lâmina é mais larga e encontra se mais fundida rostralmente ao septo interorbital. O rostro paraesfenóide articula se ventralmente com o pterigóide (facies articularis pterygoidea)e com o palatino (facies articularis palatina).Em sua extremidade rostral, na facies articularis vomeralis, articula se com o vômer. Na face ventrocaudal do rostro paraesfenóide (basisrostri parasphenoidale),observa se uma depressão (tuba aditiva communis)em forma triangular, recoberta pela projeção paraesfenóide, com seus respectivos foramens laterais.
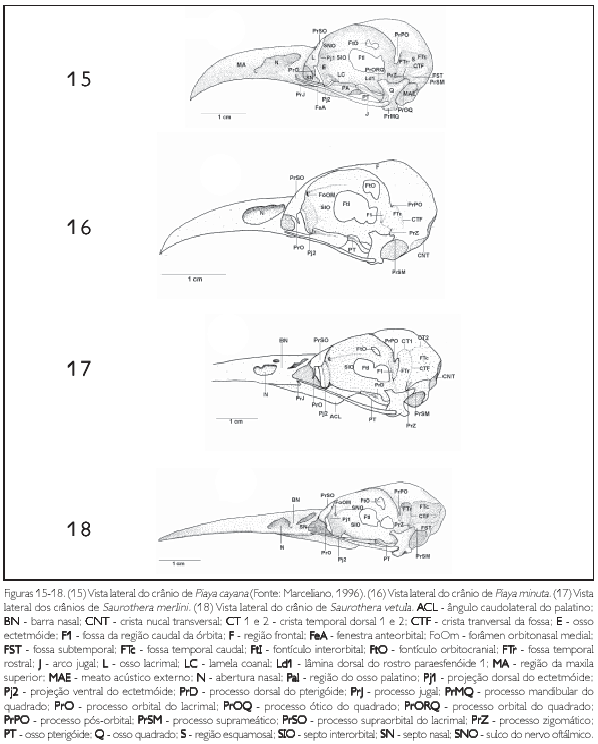
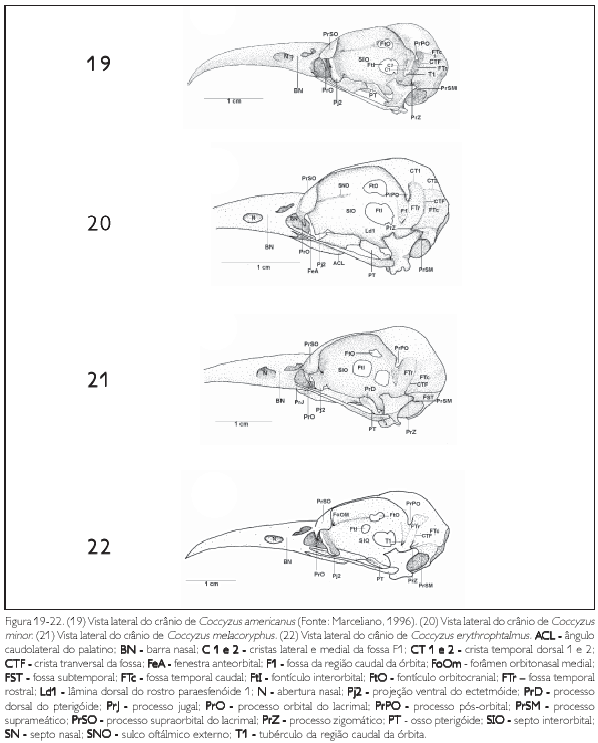
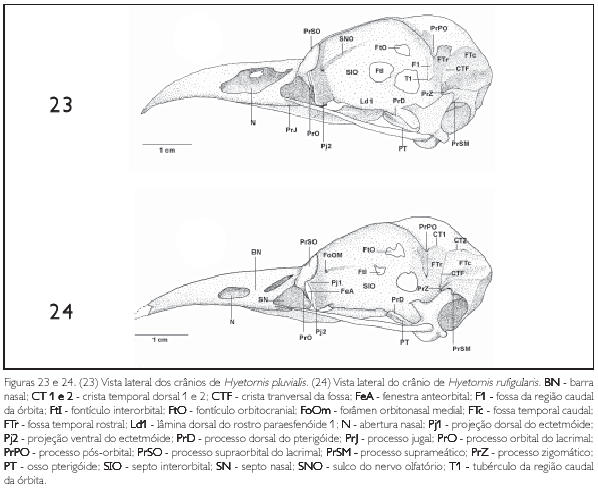
A região do osso ectetmóide (os ectethmoidale) (E: Figura 15) forma a parede rostrolateral da órbita. Sua porção dorsal (facies nasalis) funde-se à região frontal, através da projeção dorsal 1 (Pj1: Figuras 15, 18 e 24), e medialmente com a região do osso mesetmóide. Em todas as espécies esta projeção é larga lateralmente, contatando-se com a face medial do processo supra-orbital do lacrimal. A porção ventral do ectetmóide (facies orbitalis) estende-se lateroventralmente através da projeção óssea 2 (Pj2: Figuras 15-18, 19-22, 23, 24 25-28, 29-32, 33 e 34) com forma laminar que recobre parte do osso palatino, aproximando-se do arco jugal. Esta projeção possui padrão de desenvolvimento e forma semelhantes em P. cayana (Figura 15), P. minuta (Fgura 16), S. vetula (Figura 17) e S. merlini (Figura 18), nas quais se apresenta como uma fina lâmina, relativamente mais expandida lateralmente, levemente inclinada ventralmente com ápice ventral arredondado. Já em C. americanus (Figura 19). C. minor (Figura 20), C. melacorhyphus (Figura 21), C. erythrophtalmus (Figura 22), H. pluvialis e H. rufigularis (Figuras 23 e 24) a projeção ventral do ectetmóide é mais larga lateralmente (longa), ultrapassando a linha lateral do lacrimal e atingindo dorsalmente a linha lateral da barra jugal, porém, não se contata com esta última. H. rufigularis apresenta o osso uncinado (os uncinatum) (U: Figura 34) localizado ventralmente ao ectetmóide e caudalmente ao lacrimal. Este osso liga-se ao ectetmóide, arco jugal e septo nasal através de um fino tecido conjuntivo. Em H. pluvialis (Figura 33) não foi possível observar tal estrutura, uma vez que a região entre o lacrimal e ectetmóide encontra-se danificada no espécime obtido.
O lacrimal (os lacnmale) (L: Figura 15) apresenta duas porções: processo supra-orbital (proc. supraorbitalis) (PrSO: Figuras 15-18, 19-22, 23 e 24) e processo orbital (proc. Orbitalis) (PrO: Figuras 15 a 24). O primeiro limita-se rostromedialmente com a região do osso nasal através da sutura lacnmonasal (SLN: Figuras 1, 5 e 9) e o segundo rostralmente com afenestra anteorbital (fenestra antorbitalis) (FeA: Figuras 15 e 20). Em todas as espécies o processo supra-orbital é delgado, arredondado, amplo e estende-se lateralmente em relação à face lateral da região frontal. Já o processo orbital é pouco desenvolvido, espicular, com uma pequena projeção também espicular que se estende medialmente no crânio. Todavia, em S. vetula (Figura 18) é extremamente reduzido, possuindo desenvolvimento semelhante â reduzida projeção medial deste processo.
A região esquamosal (os squamosum) (S: Figura 15) é delimitada ventralmente pela face dorsal do meato acústico externo (meatus acusticus externus) (MAE: Figura 15), rostromedialmente contata-se com a região caudal da órbita por meio da crista caudal orbital 1, caudalmente com a crista nucal transversa e rostrodorsalmente com a região parietal através do processo pós-orbital. A porção ventral da região esquamosal projeta-se rostroventralmente, formando o processo zigomático (proc. zygomaticus) (PrZ: Figuras 15-18, 19-22, 23 e 24) em forma laminar achatada dorsoventralmente. Este processo é curto, pois seu comprimento varia entre 1 /4 e 1 /5 da distância entre a sua inserção no crânio e arco jugal, é curvado ventralmente, com ápice arredondado e apresenta um sulco mediano em sua face dorsal. A região esquamosal forma, ainda, o processo suprameático (proc. suprameaticus) (PrSM: Figuras 15 a 24) pouco desenvolvido e localizado ventrocaudalmente em relação ao processo zigomático; projeta-se lateroventralmente articulando-se com a face dorsal do capítulo esquamosal do processo ótico do osso quadrado. P. minuta difere das demais espécies por apresentar este processo mais longo e espicular. Todas as espécies estudadas apresentam uma crista transversal (CTF: Figuras 15 a 24) que separa a fossa temporal (fossa temporalis) em duas fossas conspícuas e profundas na região esquamosal, a saber: fossa temporal rostral (FTr: Figuras 15 a 24), delimitada pela crista temporal dorsal T1 (CT1: Figuras 17, 20 e 24) situada caudalmente ao processo pós-orbital e rostralmente limitada com a crista 1 da região caudal da órbita, encontra-se mais larga (dorsoventralmente) que comprida (rostrocaudalmente) em todas as espécies, porém, é mais profunda em H. pluvialis e H. rufigularis (Figuras 23 e 24): e fossa temporal caudal (FTc: Figuras 15, 17 a 24) situada caudalmente à fossa temporal 1, mais comprida que larga, estendendo-se à região caudal do crânio, é delimitada pela crista temporal dorsal 2 (CT2: Figuras 17, 20 e 24) laterodorsalmente e pela crista nucal transversa lateroventralmente. A crista transversal encontra-se discretamente mais proeminente em C. americanus (Figura 19), C. minor (Figura 20), C. melacoryphus (Figura. 21), C. erythropthalmus (Figura 22) que nas demais espécies, com exceção de H. pluvialis (Figura 23) e H. rufigularis (Figura 24), que apresentam a crista transversal saliente, resultado da maior profundidade da fossa temporal rostral destas espécies em relação às demais. Em P. minuta esta crista é relativamente pouco proeminente. A fossa temporal caudal encontra-se profunda e estendida caudalmente até a linha mediana, próximo à proeminência cerebelar em todas as espécies, sendo que em H. pluvialis e H. rufigularis (Figuras 9 e 10) tal fossa é mais larga que nas demais espécies. P. minuta (Figuras 2 e 16) difere das demais por apresentar a fossa temporal caudal extremamente rasa e suas respectivas cristas mconspícuas, o que dificulta inferências sobre o tamanho relativo desta fossa no espécime estudado. A fossa subtemporal (fossa subtemporalis) (FST: Figuras 15, 18 e 21) é pouco ampla e rasa se comparada à fossa temporal e em P. minuta é mais rasa, larga e mais curta que nas demais espécies.
O processo pós-orbital (proc. postorbitalis) (PrPO: Figuras 11-14, 15-18, 19-22. 23 e 24) é laminar, achatado rostrocaudalmente e estende-se em direção à porção ventral do crânio. Apresenta-se relativamente pouco desenvolvido e seu comprimento varia de 1/4 a 1/5 da distância entre sua origem no crânio até o arco jugal, exceto em P. minuta, onde este processo é extremamente reduzido, em forma de um tubérculo.
Orbita
A porção caudal da órbita (paries caudalis orbrtae) é formada pela fusão de vários ossos, funde-se caudolateralmente com a região esquamosal, dorsocaudalmente com a região parietal e medialmente limita-se com o fontículo orbitocranial e forâmen óptico. Na area muscularis aspera observa-se uma fossa 1 laterocaudoventral (F1: Figuras 16, 17, 20 e 23), delimitada lateralmente pela crista 1 lateral (C1: Fgura 19) e medialmente por uma crista 2 medial (C2: Figura 19). Esta fossa é pouco profunda e larga dorsalmente, mas se estreita ventralmente. As cristas unem-se ventralmente em um pouco proeminente tubérculo orbital 1 (T1: Figuras 19, 22 e 23). O forâmen do nervo maxilomandibular (foramen nervus maxillomandibularis) é conspícuo e, por se localizar medialmente em relação à face timpânica (facies tympanica) do quadrado, lateroventralmente na região lateroesfenóide, não pode ser observado em vista lateral. Todas as espécies descritas apresentam este forâmen com aberturas de diâmetros relativamente semelhantes.
Na paries medialis orbitae, observa-se o septo interorbital (septum interorbitale) (SIO: Figuras 15-18, 19-22, 23 e 24) delgado e laminar. Em sua região mediana observam-se as seguintes aberturas: fontículo orbitocranial (fonticuli orbitocraniales) (FtO: Figuras 15 a 24) localizado dorsomedialmente, com praticamente o mesmo diâmetro de abertura entre as espécies estudadas; forâmen óptico (foramen opticum) localizado ventromedialmente, com também aberturas semelhantes entre as espécies estudadas, e comunicam a órbita com o interior da caixa craniana, tal qual a primeira abertura citada; e fontículo mterorbital (fonticuli interorbitales) (Ftl: Figuras 15 a 24), uma abertura formada na porção central entre a lâmina dorsal 1 do rostro paraesfenóide e o septo interorbital. Nota-se que o tamanho da abertura deste fontículo depende do nível de ossificação do septo mterorbital com a lâmina dorsal do rostro paraesfenóide. Nas espécies pertencentes aos gêneros Coccyzus (Figuras 19 a 22) e Hyetornis (Figuras 23 e 24) esta abertura é menor que nas demais espécies, pois nestas últimas abrange a maior parte da região mediana do septo interorbital.
Na panes rostralis orbitae observam-se: o sulco do nervo olfatório (sulcus n. olfactorii) (SNO: Figuras 15, 18 e 23) pouco profundo e forâmen orbitonasal medial (foramen orbitonasalis medialis) (FoOm: Figuras 16, 18, 22 e 24) disposto dorsomedialmente, próximo ao septo interorbital. Este forâmen encontra-se mais desenvolvido (abertura com maior calibre) apenas em P. minuta (Figura 16) e H. pluvialis (Figura 23). Lateralmente a este forâmen, constata-se o pouco conspícuo forâmen orbitonasal lateral (foramen orbitonasalis lateralis) em todas as espécies estudadas.
Facies
Ossa maxillae
A maxila superior (ossa maxillae) (MA: Figuras 1, 15 e 25) é formada pela fusão dos ossos nasal (nasale), pré-maxilar (premaxillare) e maxilar (maxillare). Seu ápice rostral apresenta curvatura ventral nas espécies descritas, porém, nas espécies de Saurothera (Figura 18) esta curvatura é menos acentuada. A maxila superior funde-se ventrocaudalmente ao osso palatino e laterocaudalmente ao arco jugal. E estreita e longa, correspondendo há cerca de metade (50%) do comprimento total do crânio, sendo ainda mais longa e estreita em S. merlini e S. vetula (Figuras 3 e 4), que possui cerca de 3/5 (60%) do comprimento total do crânio.
Na região do osso nasal, a abertura nasal externa (apertura nasi ossea) (N: Figuras 1-4, 5-8, 9, 10, 15-18, 19-22, 23 e 24) situa-se na porção laterocaudal da maxila superior. Todas as espécies possuem o comprimento (rostrocaudal) da abertura nasal maior que sua largura (dorsoventral), estendendo-se à porção caudal da maxila superior. Todavia, nas espécies de Coccyzus (Figuras 5-8, 19-22), Saurothera (Figuras 3, 4, 17 e 18) e Hyetornis rufigularis (Figuras 10 e 24) constata-se uma lâmina óssea que abrange uma área central, formando uma pequena barra óssea nasal (BN: Figuras 3-4, 5-8, 10, 17-18, 19-22 e 24) que atravessa a abertura nasal, dividindo-a em duas partes com extensões equivalentes. Todavia, em H. pluvialis (Figuras 9 e 23) esta região encontra-se danificada, não podendo constatar a existência desta barra. Em P. cayana (Figuras 1 e 15) e P. minuta (Figuras 2 e 16) esta lâmina é ausente, possibilitando total abertura externa à narina. Rostralmente em relação ao ectetmóide e dorsalmente ao processo maxilopalatino, observa-se o septo nasal (septum nasi osseum) (SN: Figuras 15, 18, 20 e 24) achatado lateralmente, projetando-se caudalmente nafenestra anteorbital. Constata-se total fusão dos ossos nasais com os demais ossos da maxila superior, embora em alguns espécimes, na pila supranasal, observa-se uma sutura pouco conspícua, evidenciando a porção dorsocaudal do processo frontal (proc. frontalis) do nasal. Todas as espécies apresentam a face dorsal da pila supranasal (pila supranasalis) (PSN: Figura 1) plana.
Na face ventral da maxila superior os ossos maxilares encontram-se quase que totalmente fundidos entre si em todas as espécies estudadas, observando-se somente uma pequena fenestra central em alguns espécimes. Na região do osso maxilar, na porção laterocaudal da maxila superior, observa-se o processo jugal (proc. jugalis) (PrJ: Figuras 15, 21, 25 e 34) achatado lateralmente e disposto ventralmente ao arco jugal.
Na porção ventrocaudal da maxila superior, rostromedialmente em relação ao palato, destacam-se os processos maxilopalatinos (processum maxillopalatinum) (PrMP: Figuras 25-28, 29-32, 33 e 34) bem desenvolvidos, longos, fundidos medialmente, somente não se fundindo entre si na porção caudal. Expandem-se dorsalmente no crânio e lateroventralmente em relação ao septo nasal em todos os Coccyzinae estudados.
Palati
A região do palato é compreendida pelos ossos palatino (os palatinum), vômer (os vomer) e pterigóide (os pterygoideum).
O osso palatino (PA: Figuras 15 e 25) dispõe-se como uma lâmina posicionada horizontalmente; funde-se rostralmente à maxila superior, na zona flexoria palatina, através do processo maxilar (proc. maxillaris) (PrM: Figuras 25-28, 29-32, 33 e 34) mais estreito rostralmente em relação à porção caudal do palatino. O palatino expande-se lateralmente, formando a conspícua crista lateral (crista lateralis) (CL: Figuras 25-28, 29-32, 33 e 34), que apresenta leve curvatura ventral. Esta crista estende-se até a porção caudal do palatino, terminando de forma abrupta (perpendicular) nos limites com o processo pterigóideo do palatino e forma o ângulo caudolateral (angulus caudolateralis) (ACL: Figuras 25 a 34). A porção laterocaudal do palatino apresenta um pequeno processo, sendo que nas espécies de Saurothera (Figuras 27 e 28) é mais longo que nas demais espécies. Todavia, em muitos espécimes analisados a porção laterocaudal do palatino encontra-se danificada e nada se pode inferir em relação a este processo nestes espécimes. Em P. minuta a porção caudal do palato é reduzido se comparada às demais espécies.
A fossa coanal (fossa choanalis) (FC: Figura 25) está delimitada lateroventralmente pela crista ventral (crista ventralis) (CV: Figuras 25-28, 29-32, 33 e 34) e dorsalmente pela lamela coanal. Essa fossa é rasa e estreita, estreitando-se ainda mais caudalmente devido à aproximação medial dos processos pterigóideos do palato. Dentre estas espécies, P. minuta (Figura 26) apresenta essa fossa relativamente mais larga. Entre as cristas lateral e ventral observa-se a fossa ventral (fossa ventralis) (FV: Figura 25) rasa e larga (ampla). A crista ventral é estreita, pontiaguda rostralmente, sendo que em P. minuta (Figura 26) é menos proeminente.
O palato curva-se dorsalmente próximo à extremidade rostral do paraesfenóide, formando a lamela coanal (lamela choanalis) (LC: Figura 15), que é curta e pouco profunda nas espécies estudadas, restringindo-se à porção rostral da face dorsal do osso palatino. Os processos pterigóideos do palatino (proc. pterygoideum) (PrPT: Figuras 25-28, 29-32, 33 e 34) são pouco desenvolvidos. Os processos maxilopalatmos da maxila superior, por serem longos e fundidos entre si, ocupam a maior parte da área entre os processos maxilares, caracterizando um palato do tipo fechado.
Na maioria dos espécimes estudados o vômer está ausente, porém, quando presente, situa-se na região dorsorostral da fossa coanal e medial aos processos maxilopalatinos, articulando-se caudalmente com a extremidade rostral do rostro paraesfenóide e ventralmente com a facies articularis palatina. Apresenta-se reduzido e afilado rostralmente e não se contata com o septo nasal.
O pterigóide (PT: Figuras 25-28, 29-32, 33 e 34) é estreito e longo, afila-se em sua face dorsal emitindo uma bem desenvolvida lâmina denominada processo dorsal (proc. dorsalis) (PrD: Figuras 17, 21, 23 e 24), porém, em H. rufigularis (Figura 24) o processo dorsal é pouco proeminente quando comparado ao das demais espécies. Na facies articularis quadratica o pterigóide apresenta uma cótila (CoPt: Figura 14) que se articula com o côndilo pterigóideo do osso quadrado e na facies articularis palatina articula-se com palatino através de uma reduzida expansão rostrolateral (pars palatina) (PP: Figuras 28 e 34).
O arco jugal (arcus jugalis) (J: Figuras 1, 15 e 25) é formado pela fusão dos ossos jugal (os jugalis) e quadradojugal (os quadratojugalis). Trata-se de uma barra óssea que articula a maxila superior ao osso quadrado. Funde-se rostralmente à face dorsal do processo jugal da maxila superior e na porção caudal articula-se, através do côndilo quadradojugal (condylus quadratojugalis), com a cótila quadradojugal (cotyla quadratojugalis) do côndilo lateral do processo mandibular do osso quadrado. O arco jugal é comprido, estreito, alargando-se medialmente em sua porção rostral (Figura 25) e levemente inclinado dorsoventralmente em relação à caixa craniana.
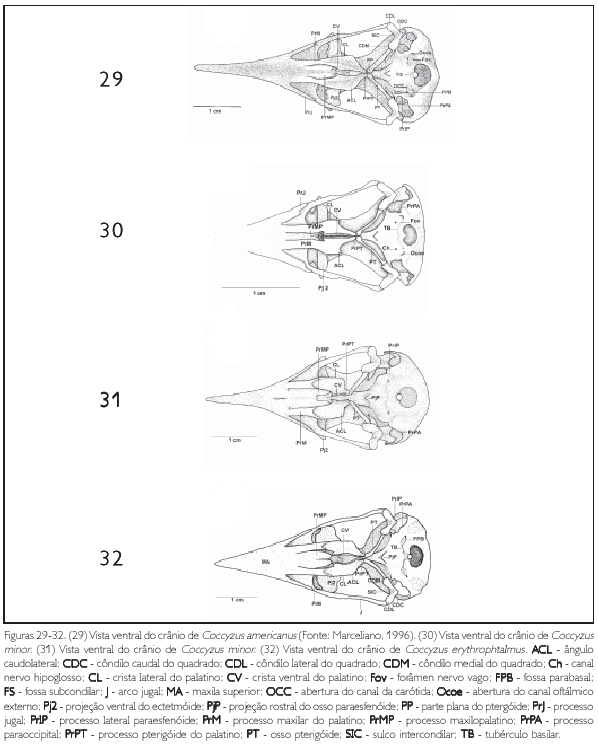
O osso quadrado (os quadratum) (Q: Figuras 11 e 25) apresenta uma parte central, denominada corpo do quadrado (corpus ossi quadrati). Contém três processos: 1) processo ótico (proc. oticus) (PrOQ: Figuras 15 e 25) estende-se dorsocaudalmente a partir do corpo do quadrado, apresentando uma base alargada, estreitando-se até a sua superfície de articulação com a região esquamosal, onde se bifurca e apresenta duas superfícies de articulação, a) capítulo esquamosal (capitulum squamosum) (CAS: Figura 25) que se articula com a face ventrocaudal do processo zigomático e com a face rostroventral do processo suprameático e b) capítulo ótico (capitulum oticum) (CAO: Figura 25), que se articula com a face ventral da região dorsal do meato acústico externo, sendo mais desenvolvido que o capítulo esquamosal; 2) processo orbital (proc. orbitalis) (PrORQ: Figura 15) é achatado, curto e largo, projeta-se obliquamente na órbita em um ápice pontiagudo rostralmente, sendo ainda mais curto em P. minuta e nas espécies de Coccyzus; 3) e processo mandibular (proc. mandibularis) (PrMQ: Figura 15), que se localiza na porção ventral do corpo do quadrado, articulando-se com a mandíbula e pterigóide e apresentando quatro côndilos: lateral, medial, caudal e pterigóideo (condylus lateralis, medialis, caudalis e pterygoideus). Os côndilos lateral (CDL: Figura 29) e medial (CDM: Figura 29) apresentam praticamente o mesmo desenvolvimento, porém, o côndilo medial afila-se ventralmente, enquanto o lateral é arredondado. O côndilo caudal (CDC: Figura 29) é reduzido em relação aos demais côndilos supracitados. O côndilo lateral apresenta pequena depressão em sua porção laterodorsal (cotyla quadratojugalis), local de articulação com o arco jugal. Ainda neste processo observa-se, entre os côndilos lateral, medial e caudal, o sulco mtercondilar (sulcus intercondylaris) (SIC: Figura 29) estreito e profundo. O côndilo pterigóideo é o côndilo mais reduzido deste processo, localiza-se na face medial do corpo do quadrado, articulando-se com a cótila caudal do osso pterigóide. Na face medial do quadrado (facies tympanica) constata-se uma depressão (sulcus pneumaticum) rasa, limitada pela crista timpânica (crista tympanica) também pouco conspícua. Em termos gerais, o osso quadrado apresenta a mesma forma entre as espécies de Coccyzinae.
Ossa mandibulae
A mandíbula das aves é formada pelos ossos articular (os articulare),angular (os angulare),supra-angular (os suprangulare),dental (os dentale),esplenial (os spleniale),pré-articular (os prearticulare)e, em alguns grupos em particular, também pelo coronóide (os coronoideum)fundidos. Pode ser subdividida em parte sinfisial (pars symphysialis) (PSI: Figura 35), parte intermediária (pars intermedia) (PI: Figura 35) e parte caudal (pars caudalis) (PC: Figura 35). Quanto às dimensões da mandíbula, percebe-se que é longa e estreita, sendo o comprimento total cerca de 2 a 3 vezes maior que a distância entre os dois ramos mandibulares (ramus mandibulae),que são estreitos e delgados.

A parte sinfisial reúne os dois ramos da mandíbula e atinge cerca de 1/3 do comprimento total da mandíbula em P. cayana (Figura 35), P. minuta, S. merlini (Figura 36), S. vetula, C. americanus (Figura 37), C. minor, C. melacoryphus e C. erythropthalmus. Entretanto, em H. pluvialis e H. rufigularis (Figura 38) a parte sinfisial é menos desenvolvida, pois atinge somente cerca de 1/6 desta medida.
O osso dental é a principal porção de cada ramo mandibular e corresponde em sua maioria à porção dorsal e lateral da parte intermediária da mandíbula. Em sua face dorsal, observa-se o ângulo mandibular (angulus mandibulae) (AM: Figura 39) inclinado ventralmente e levemente expandido lateralmente. Todavia, nas espécies de Saurothera este ângulo é discretamente inclinado (praticamente reto).
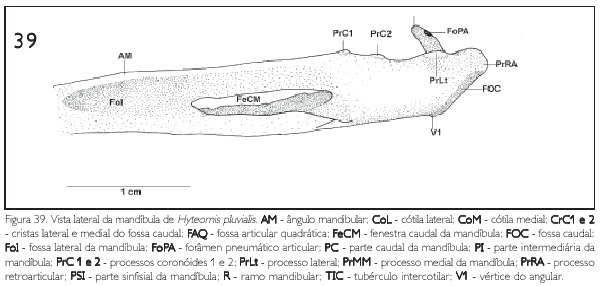
Na face dorsal da parte caudal da mandíbula, os processos coronóideos 1 e 2 (proc. coronoideum) (PrCl e PrC2: Figura 39) são pouco desenvolvidos e com desenvolvimento semelhantes em todas as espécies aqui estudadas.
Na face medial observa-se o tubérculo pseudotemporal (tuberculum pseudotemporalis) reduzido e pouco conspícuo em todas as espécies analisadas. Ainda nesta face, a fenestra caudal (fenestra caudalis) (FeCM: Figura 39) encontra-se desenvolvida, porém em P. cayana e Rminuta observa-se uma lâmina óssea, um fino tegumento que recobre toda a fenestra. Já nas demais espécies observa-se tal lâmina, porém, pouco recobre a fenestra caudal (Figura 39). As espécies estudadas não apresentaram a fenestra rostral (fenestra rostralis).Constata-se uma depressão na face medial da mandíbula (fossa aditus canalis mandibulae) rasa e ampla. Já na face lateral, a fossa lateral (fossa lateralis) (Fol: Figura 39) é profunda, principalmente em sua porção rostral, e ampla.
A face dorsal da parte caudal da mandíbula caracteriza-se por ser o local de articulação com o crânio, destacando-se a fossa articular (fossa articularis quadrática) (FAQ: Figuras 35-38) profunda e larga, delimitada lateralmente pela cótila lateral (cotyla lateralis) (CoL: Figura 35) e medialmente pela cótila medial (cotyla medialis) (CoM: Figura 35), sendo estas cótilas semelhante entre si quanto à extensão. Já a cótila medial é relativamente mais profunda que a lateral para abrigar o côndilo medial do quadrado, que apresenta sua extremidade ventral afilada. Entre as cótilas destaca-se o proeminente tubérculo mtercotilar (tuberculum intercotylaris) (71C: Figuras 35-38 e 40). Ainda nesta região observa-se o processo medial (proc. medialis mandibulae) (PrMM: Figuras 35-38, 39 e 40) desenvolvido, comprido, espesso na base, afilando-se dorsalmente e projetando-se dorsomedialmente em direção ao crânio. Este processo apresenta, em sua porção lateral, o forâmen pneumático (foramen pneumaticum articulare) (FoPA: Figuras 38 a 40). Na face lateral da região articular da mandíbula observa-se pouco desenvolvido processo lateral (proc. lateralis) (PrLt: Figuras 35-38, 39 e 40) com o desenvolvimento semelhante entre as espécies consideradas. Na extremidade caudal da mandíbula observa-se o processo retroarticular (proc. retroarticularis) (PrRA: Figuras 35-38, 39 e 40) curto e pouco desenvolvido.

Na face ventrocaudal da parte caudal da mandíbula encontra-se a fossa caudal (fossa caudalis) (FOC: Figuras 39 e 40) rasa e estreita, limitada pelas cristas 1 (lateral) e 2 (medial) (CrCI e CrC2: Figura 40) pouco proeminentes. Ventralmente estas duas cristas unem-se, formando o reduzido vértice 1 do angular (VI: Figura 39).
DISCUSSÃO
Considerações osteológicas
A anatomia foi um tema bastante explorado no final do século XIX e, embora tenha contribuído grandemente para a compreensão dos grandes grupos de Aves, os estudos anatômicos foram pouco elucidativos sistematicamente em níveis categóricos inferiores, pois muitas vezes a amostragem era inadequada e freqüentemente as comparações eram feitas entre táxons filogeneticamente distantes entre si. Contudo, com o advento da sistemática filogenética, notou-se um aumento de estudos anatômicos na classe Aves nas últimas décadas. Em relação aos Cuculiformes, nas últimas décadas, destacam-se os trabalhos de Seibel (1988), Hughes (2000), Posso (1999) e Posso e Donatelli (2001), que utilizaram caracteres osteológicos para análise cladística, e de Marceliano (1996), que estudou a miologia e osteología craniana das espécies Crotophaga ani, Guira guira, Coccyzus americanus e Piaya cayana.
Antes do estabelecimento das relações osteológicas entre as espécies de Coccyzinae, é importante discutir e comparar determinadas variações morfológicas com outros cucos e demais ordens, procurando interpretá-las em um contexto geral, visando a contribuir para o refinamento da osteología nos Cuculiformes, particularmente nos Coccyzinae.
Dois aspectos são importantes para discussão do ectetmóide: o desenvolvimento de sua projeção óssea ventral e sua fusão com o mesetmóide. A luz dos resultados, percebe-se que o desenvolvimento da projeção óssea ventral do ectetmóide constitui-se em um parâmetro relevante para diferenciação das espécies de Hyetornis das demais (Tabela 1). Em relação ao segundo aspecto, possivelmente o maior desenvolvimento do forâmen orbitonasal medial encontrado em P. minuta e H. pluvialis deva-se às diferenças no padrão de desenvolvimento e ossificação da projeção dorsal do ectetmóide com o mesetmóide, fato também observado por Pycraft (1903). Como exemplo, podem ser citadas as espécies de Crotophaga e em Neomorphus geoffroyi (POSSO, 1999; POSSO, DONATELLI, 2001), nas quais o processo dorsal do ectetmóide é mais curto, encontrando-se parcialmente fundido com a região do mesetmóide, o que proporciona uma maior abertura ao forâmen orbitonasal mediai como em P. minuta e H. pluvialis. Há, ainda, a possibilidade de que o tamanho deste foramen dependa do estágio ontogenético em que o semaforonte encontra-se e não propriamente trata-se de uma modificação de origem genealógica, o que dificulta seu uso em análises cladísticas.
Thompson (1899) e Jollie (1957) denominam o osso lacrimal de pré-frontal (correspondendo ao pré-frontal dos Reptilia) e este último afirma que o pré-frontal das Aves é confundido com o lacrimal dos Synapsida. Atualmente, a maioria dos autores utiliza o termo lacrimal, seguindo a orientação de Cracraft (1968) e o referencial da Nomina Anatomica Avium (BAUMEL et al., WITMER, 1993). Em termos gerais, a descrição do lacrimal corrobora a de Huxley (1868), Shufeldt (1901), Pycraft (1903) e Marceliano (1996), embora Pycraft (1993) afirme que, contrariamente ao descrito neste trabalho, o lacrimal não se contata com o osso nasal nos Cuculiformes.
Segundo Baumel e Witmer (1993), o osso uncmado é observado nos Musophagiformes (turacos, aves endêmicas da Africa), o que foi constatado na observação de algumas espécies pertencentes a este grupo (obs. pess.) em alguns exemplares do Museu de História Nartural de Taubaté (MHNT). Lowe (1943) afirma que os cucos não apresentam o osso uncinado, mas este autor analisa somente alguns gêneros desta ordem. Já Pycraft (1903) afirma que os cucos podem apresentar o osso uncinado. Esta estrutura foi observada em Scythrops, Eudynamis e Carpococcyx radiceus (Cuculidae) por Beddard (1885, 1901). Além disso, pode-se aqui observar tal estrutura em Hyetornis rufigularis (Figura 34). Embora esta estrutura acessória craniana ocorra também em alguns grupos de aves filogeneticamente distantes dos cucos e turacos (BURTON, 1970), a existência desta estrutura nos cucos e turacos pode corroborar um parentesco entre eles, o que é aceito por diversos autores, incluindo trabalhos sistemáticos atuais (SEIBEL, 1988; HUGHES, 2000), contrariamente à hipótese de Sibley e Ahlquist (1990), os quais propõem que os cucos e os turacos sejam grupos distantes filogeneticamente.
Na descrição da região esquamosal, Pycraft (1903) observou que todas espécies pertencentes à família Musophagidae (turacos) possuem a fossa temporal confinada à parte lateral da caixa craniana, sendo pouco conspícua (rasa); enquanto nos cucos esta fossa estende-se à parte caudal, quase atingindo a linha mediana, sendo bastante conspícua e profunda.
Esta conformação de fossa foi observada nos Coccyzinae aqui estudados, principalmente no que se refere às espécies congêneres de Hyetornis. A única exceção é em Piaya minuta, que apresenta esta fossa pouco desenvolvida. A fossa temporal, praticamente conspícua em Piaya minuta, pode ser explicada pelo pequeno tamanho do crânio (miniaturização) apresentado por esta espécie, causando dificuldades para observação destas estruturas de forma conspícua.
A profundidade da fossa temporal depende do desenvolvimento do sistema adutor mandibular externo, ou seja, quanto mais desenvolvido este sistema muscular maior a profundidade da fossa temporal (MORIOKA, 1974), também observada nos Picidae (DONATELLI, 1996). Tal característica pode ser atribuída também aos Coccyzinae e Crotophaginae, já que Marceliano (1996) observou este sistema muscular desenvolvido, juntamente com a fossa temporal profunda, em C. americanus, G. guira e em C. ani. São necessários estudos miológicos nos demais Cuculiformes para que seja possível observar se esta relação ocorre de maneira geral nos cucos.
A crista transversal que divide a fossa temporal, embora evidente, curiosamente foi pouco descrita na literatura, com exceção do trabalho de Marceliano (1996). Tal crista parece ocorrer de maneira geral nos cucos (POSSO, 1999), podendo se constituir em urna importante estrutura a ser utilizada em análise filogenética.
O processo zigomático é denominado processo esquamosal na maioria dos trabalhos consultados e processo zigomático por Baumel e Witmer (1993) e Zusi (1993). Embora aqui não se concorde com a denominação processus zygomaticus, por lembrar uma estrutura característica dos Mammalia (Synapsida), utiliza-se tal nomenclatura, já que se propôs seguir Baumel e Witmer (1993).
A parede caudal da órbita, em geral, é formada pela fusão dos ossos frontal (os frontale), orbitoesfenóide (os orbitosphenoidale), pleuroesfenóide (os pleurosphenoidale) e lateroesfenóide (os laterosphenoidale), o que torna difícil delimitaríais ossos sem um estudo ontogenético. Marceliano et al. (manuscrito) observaram que, em um indivíduo jovem de Penelope superciliaris (Cracidae, Galliformes), o osso frontal e orbitoesfenóide participam da maior parte da parede caudal da órbita, assim como observado em Gallus gallus (Phasianidae, Galliformes) por Jollie (1957). Ladeira (1997), estudando Bucconidae, considera esta região como lateroesfenóide; Zusi (1993) como região pleuroesfenóide; e Höfling (1995), estudando Ramphastidae e Donatelli (1992, 1996, 1997), estudando Piciformes e Dendrocolaptidae (Passeriformes), consideram esta região como orbitoesfenóide. Tendo em vista que não há trabalhos ontogenéticos que indiquem as respectivas contribuições de cada osso nos Cuculiformes, esta porção da órbita foi denominada neste trabalho como parte caudal da órbita.
No que se refere à maxila superior, nota-se que esta região foi descrita detalhadamente por Pycraft (1903). Em termos gerais, a descrição do presente trabalho concorda com a realizada por este autor. Por haver diferentes níveis de ossificação do osso nasal com os ossos pré-maxilar e maxilar, a forma das narinas externas está fortemente modificada nos Cuculidae (BEDDARD, 1885; PYCRAFT, 1903). Isto pode ser verificado na descrição das espécies estudadas. Assim, nas espécies de Piaya as narinas possuem sua abertura externa livre, enquanto nas espécies de Coccyzus, Saurothera e Hyetornis rufigularis constatou-se uma fina lâmina óssea que recobre a área central da narina, dividindo-a em duas aberturas (Tabela 1). Em relação aos demais Cuculiformes, segundo Shufeldt (1901) e Posso e Donatelli (2001) em Crotophaga (Crotophagidae), as narinas estão situadas relativamente em posição lateral e sua abertura externa é menor e circular. Beddard (1885) observou um padrão de abertura nasal semelhante ao de Crotophaga em Scythrops (Cuculidae). Em Neomorphus, Morococcyx e Geococcyx (Neomorphmae) somente a porção rostral da abertura nasal não é obstruída por uma lâmina nasal (POSSO, 1999).
Ainda em relação à região nasal observa-se uma condição holorrma (GARROD. 1873) nos Coccyzinae, uma vez que os ossos nasais encontram-se fundidos aos demais ossos da maxila superior, embora em alguns espécimes seja possível observar uma sutura pouco conspícua na região da pila supranasal. Geralmente a condição holorrina está associada ao procmetismo nas Aves (BOCK, 1964; ZUSI, 1984), concordando com as observações feitas nas espécies aqui descritas que apresentam o osso mesetmóide não associado ao septo nasal (condição procmética).
A região do palato constitui-se em uma das mais importantes regiões para diferenciação dos Cuculiformes dos Musophagiformes, pois nestes os palatinos estão separados em sua porção caudoventral, sendo possível observar a porção rostral do paraesfenóide; nos Cuculiformes os palatinos estão juntos nesta posição, corroborando a descrição de Pycraft (1903). No presente trabalho observou-se que a região do palato apresenta praticamente o mesmo padrão entre as espécies aqui analisadas e a descrição desta região é semelhante àquelas observadas por outros autores (HUXLEY, 1867; BEDDARD, 1885; SHUFELDT 1901; PYCRAFT, 1903; MARCELIANO. 1996), principalmente no que se refere à condição desmognata do palato (HUXLEY 1868), uma vez que o palato é fechado e o vômer é vestigial ou inexistente. Todavia, foram observadas variações em relação ao processo laterocaudal e fossa ventral (Tabela 1).
No que se refere ao processo laterocaudal do palatino, observado principalmente no gênero Saurothera, Feduccia (1973) constatou um processo nesta mesma posição em Furnariidae e Dendrocolaptidae (Passeriformes) e o denominou de 'Transpalatine Process', processo também observado em alguns gêneros de Dendrocolaptidae por Donatelli (1997). Por ser bastante reduzido nos Coccyzinae, não se denominou aqui tal processo como sugere Feduccia (1973), pois não ultrapassa o palatino como está explícito no nome supracitado.
A região do osso pterigóide nos Cuculidae estudados difere consideravelmente da dos Musophagidae nos quais este é relativamente menor, mais espesso, mostrando vestígios da face basipterigóide (PYCRAFT 1903). Segundo este autor, o pterigóide pouco varia nos cucos. Tal característica foi constatada na descrição dos Coccyzinae, já que o pterigóide apresenta pouca variação significativa, somente no que se refere ao desenvolvimento do seu processo dorsal (Tabela 1).
A região exoccipital abriga o canal do nervo hipoglosso, forâmen do nervo vago e canal oftálmico externo, conforme estudos ontogenéticos em Gallus gallus, realizados por Jollie (1957). Donatelli (1992), estudando Galbulidae; Marceliano (1996), comparando Opisthocomus hoazin com Cracidae, Musophagidae e Cuculidae; e Ladeira (1997), estudando Buccomdae, descreveram estas aberturas como pertencentes à região basioccipital. No entanto, estas aberturas são consideradas tanto da região exoccipital, como da região basioccipital, segundo Baumel e Witmer (1993). No presente trabalho optou-se por considerar estas aberturas como pertencentes à região exoccipital, já que o osso exoccipital abrange a maior parte da região occipital, sendo os ossos supra-occipital e basioccipital pequenos, estreitos e restritos à porção mediana da região occipital, segundo observações realizadas por Pycraft (1903) em indivíduos jovens de Cuculus canorus (Cuculmae) e de Geococcyx californianus (Neomorphinae).
O processo lateral paraesfenóide não varia entre os Coccyzinae e o processo medial paraesfenóide é ausente, corroborando as observações de Bock (1960).
Percebe-se, através da literatura, que existem poucas descrições da mandíbula nos Cuculiformes. Descrições da mandíbula são citadas nos trabalhos de Pycraft (1903) e Marceliano (1996), sendo que a descrição de Marceliano (op. cit.) é mais completa e pouco diverge da descrição realizada neste trabalho.
Pycraft (1903) e Marceliano (1996) descrevem o processo coronóide em Cuculiformes e Musophagidae. Segundo Parker (1890, apud Jollie 1957), o osso coronóide participa na formação da mandíbula em Passeriformes. Jollie (1957) observou este osso somente em águias (Falconiformes). Richards e Bock (1973) e Donatelli (1996) denominaram este processo de pseudocoronóide, pelo fato de não existir o osso coronóide na mandíbula das aves adultas estudadas ou por não se poder determinar, com segurança, se este osso participa na formação deste processo. Baumel e Wrtmer (1993) citam esta estrutura como processus coronoideus, mas salientam que o mesmo não está presente em todos os grupos de Aves. No que se refere aos Cuculidae, Pycraft (1903) constatou a presença do osso coronóide na mandíbula de indivíduos jovens pertencentes a Cuculus canorus (Cuculinae) e Geococcyx californianus (Neomorphinae). Por estemotivo optou-se, neste trabalho, por denominá-lo processo coronóide, embora Pycraft (1903) não tenha determinado se este osso participa da formação do processo em questão. São necessários estudos ontogenéticos nos Coccyzinae, visando a esclarecer dúvidas quanto às possíveis contribuições deste osso na formação da mandíbula.
As mandíbulas dos Coccyzinae possuem várias similaridades entre si, porém, há variações intergenéncas relevantes que separam determinados táxons (Tabela 1). Dentre as estruturas divergentes destacam-se o desenvolvimento da parte sinfisial, ângulo mandibular e fenestra caudal. As diferenças no desenvolvimento da fenestra mandibular observadas entre as espécies pode ser explicado pelos diferentes níveis de fusão entre os ossos supra-angular e dental, pois, segundo Jollie (1957), a fenestra mandibular penetra a mandíbula entre as margens de ossificação destes dois ossos. Embora esta característica separe as espécies de Piaya dos demais Coccyzinae pode haver diferenças desta abertura entre os semaforontes intra-específicos, sendo necessários estudos ontogenéticos que comprovem ou refutem tal suposição.
E evidente a necessidade de estudos ontogenéticos nos cucos, visando a esclarecer as contribuições das estruturas ósseas na formação das regiões cranianas. Necessidade também apontada por Méndez (1998) em estudos com Alcedmidae (Coracirformes). Além disso, trajetórias ontogenéticas poderiam fornecer caracteres adicionais que possivelmente indicariam relações de parentesco entre as espécies em análises claciísticas (NELSON, 1978; KLUGE. 1985; KRAUS, 1988; WAKE, 1989; WHEELER, 1990; PINNA, 1994).
Aspectos sistemáticos
De acordo com McKitrick (1991), nas últimas duas décadas as pesquisas em classificação das Aves têm demonstrado maior preocupação em conduzir os estudos com mais rigor para que possam testar, da melhor maneira possível, a legitimidade dos caracteres com que foram baseadas as classificações tradicionais, como as de Fürbringer (1888) e Gadow (1892).
Seibel (1988) considera a osteologia pós-craniana como base para inferir parentesco nos Cuculiformes, utilizando a sistemática filogenética. Contudo, este trabalho apresenta uma classificação bastante controversa, principalmente porque alia grupos bastante divergentes e distantes geograficamente. Uma vez que muitos cucos adaptaram-se ao ambiente terrestre, e que Seibel (1988) alia estes grupos com base principalmente na anatomia da cintura pélvica, pode ter ocorrido a reunião destes grupos por convergência, já que esta estrutura, bem como seus anexos, sofrem forte influência ambiental em aves que apresentam hábitos terrestres (PAYNE, 1997). Hughes (2000) também utiliza a osteologia e sistemática filogenética nos Cuculidae. Todavia, os táxons terminais são gêneros, não havendo nenhuma alusão em relação às espécies.
É importante salientar que um dos objetivos deste trabalho é procurar obter caracteres osteológicos para serem utilizados em uma futura análise filogenética nos Cuculidae. Sendo assim, tais semelhanças abaixo discutidas não implicam necessariamente em parentesco, sendo necessários estudos cladísticos que indiquem quais semelhanças são características apomórficas compartilhadas (sinapomohias).
Após a análise dos resultados obtidos, pode-se estabelecer um padrão osteológico craniano para Coccyzinae. Todas as espécies apresentam: região orbital frontal estreita, processo supraorbital do lacrimal largo e delgado, processo orbital do lacrimal afilado, ectetmóide largo, aberturas nasais amplas, processo zigomático reduzido e sulcado dorsalmente, pós-orbital pouco desenvolvido, fossa temporal profunda e larga, processo orbital do quadrado curto e largo e parte sinfisial da mandíbula bem desenvolvida. Permitiu-se chegar a este padrão devido às espécies de Coccyzinae apresentarem a maioria das regiões do crânio similares em relação à forma e desenvolvimento das estruturas, entre as quais se destacam a região paraesfenóide e os ossos lacrimal, vômer, pterigóide e quadrado.
Por outro lado, algumas regiões apresentaram as estruturas relativamente diversificadas entre as espécies estudadas: regiões esquamosal, maxila superior e mandíbula e os ossos palatino e ectetmóide. Estes caracteres serão importantes para uma futura análise cladística para teste da validade desta subfamilia como um grupo natural (monofilético), bem como suas relações com os demais Cuculidae, uma vez que há controvérsias sobre estes aspectos entre os sistematas (PETERS, 1940: SEIBEL, 1988; SIBLEY; AHLQUIST, 1990; HUGHES, 1996, 2000; PAYNE, 1997).
Apesar da aparente homogeneidade do crânio apresentada pelas espécies de Coccyzinae, pode-se constatar algumas alterações que fornecem algumas relações de similaridade e/ou diferenças. Coccyzus parece estar mais próximo de Saurothera e Hyetornis, uma vez que se pode encontrar uma fina barra óssea que atravessa a narina externa nas espécies destes gêneros, já em Piaya e nos demais Cuculidae estudados (SHUFELDT, 1901; PYCRAFT 1903; MARCELIANO, 1996; POSSO, 1999) não há tal estrutura. Todavia, Piaya e Saurothera assemelham-se por apresentar o fontículo interorbital amplo, ao passo que Coccyzus e Hyetornis apresentam esta estrutura reduzida e, ainda, apresentam a projeção ventral do ectetmóide e a lâmina dorsal do rostroparaesfenóide mais largas que em Piaya e Saurothera.
O gênero Piaya particulariza-se dos demais por apresentar a narina ampla e um tegumento recobrindo a fenestra caudal da mandíbula. Ainda, P. minuta apresenta várias características que diferem das demais espécies de Piaya e inclusive dos demais Coccyzinae: a depressão lateral do teto craniano é mais profunda e estreita, o processo suprameático é mais logo e espicular, a fossa temporal caudal é mais curta e rasa, a crista temporal tranversal é pouco saliente, a fossa temporal é mais rasa, a fossa subtemporal é mais rasa, larga e curta, o processo pós-orbital é reduzido, a forma do processo pós-orbital é tubercular, a porção caudal do palato é mais estreita, a fossa coanal é mais larga e o processo orbital do quadrado é mais curto, sendo esta última característica semelhante a Coccyzus.
Tais modificações morfológicas citadas são significativas, porque indicam uma grande modificação no crânio de P. minuta em relação às demais espécies de Piaya, bem como em relação os demais Coccyzinae. Tais modificações podem estar indicando seu posicionamento em um gênero distinto de Piaya. Payne (1997) sugere maiores investigações taxonómicas para este gênero.
Todavia, P. minuta assemelha-se às demais espécies de Piaya em relação à extremidade distai da projeção ventral do ectetmóide, processo orbital do lacrimal e fossa caudal da mandíbula, bem como em relação à plumagem, comportamento e distribuição geográfica (SIBLEY; MONROE, 1990; HUGHES, 1996; SICK, 1997). Ainda, alguns caracteres que diferem P. minuta das demais espécies congêneres podem ser decorrentes da miniaturização do crânio de P. minuta, o que dificulta estabelecer homologias primárias seguras de determinados tamanhos relativos de certas estruturas. Como exemplo, a região da fossa temporal praticamente inconspícua em Piaya minuta pode ser explicada pelo pequeno tamanho do crânio, que dificulta alojar uma musculatura mais volumosa, causando dificuldades para comparação do tamanho relativo das estruturas existentes nesta região.
Em relação ao gênero Saurothera, sua caracterização foi de fácil diagnóstico, pois apresenta várias características particulares: curvatura frontal orbital mais acentuada, depressão lateral do teto craniano mais rasa, curvatura maxilar reduzida, maxila e processo laterocaudal do palatino mais longos e inclinação do ângulo mandibular praticamente ausente. Ainda, S. vetula distingue-se de S. merlini por apresentar o processo orbital do lacrimal reduzido.
Hyetornis caracterizou-se por apresentar: fossa temporal rostral mais profunda, crista transversal proeminente e parte sinfisial da mandíbula reduzida. H. pluvialis e H. rufigularis distinguem-se entre si da seguinte forma: em H. pluvialis o fontículo medial é amplo e em H. rufígularis o processo uncinado está presente e o processo dorsal do pterigóide é reduzido.
Coccyzus foi o gênero que menos apresentou características peculiares: a depressão frontal é mais profunda e a crista transversal possui desenvolvimento intermediário entre Hyetornis e Piaya/Saurothera. C. minor, C. americanus, C. erythrophthalmus, C. melacoryphus não apresentam variações osteológicas cranianas entre si para que pudessem ser distinguidas, exceto diferente desenvolvimento (tamanho) do crânio e das estruturas associadas, principalmente em relação a C. minor.
Em síntese, pode-se perceber que há pouca variação interespecífica congênere em relação à osteología craniana no que se refere aos Coccyzinae aqui estudados, com exceção de Piaya minuta discutida acima. Observam-se maiores diferenças em nível genérico. Ainda, é difícil estabelecer relações intergenéricas nos Coccyzinae, uma vez que os caracteres cranianos provavelmente não são suficientes para tal finalidade, embora Coccyzus aparenta estar mais próximo de Hyetornis. Todavia, as estruturas ósseas cranianas indicam similaridades entre os Coccyzinae, mas não é possível discutir aqui se tais similaridades indicam monofilia para o grupo (como em SIBLEY; AHLQUIST, 1990; PAYNE, 1997).
Faz-se necessário estudos que contemplem as espécies de Coccyzinae, bem como os demais cucos em uma análise cladística, visando a contribuir para resolver esta questão, bem como estabelecer relações genealógicas entre os táxons que compõem esta subfamilia, principalmente sobre a validade e posicionamento do táxon Piaya minuta.
AGRADECIMENTOS
Agradecimentos a Elizabeth Höfling e Márcia Cristina Pascotto, pelas excelentes críticas e sugestões anatômicas; a Mana Luiza Videira Marceliano, por permitir o uso de algumas de suas ilustrações; e aos empréstimos e confiança concedidos pelos museus: Museu Paraense Emílio Goeldi, em nome de Maria Luiza Videira Marceliano; Museum of Zoology, University of Michigan, em nome de Robert B. Payne; United States National Museum, Smithsonian Institution, em nome de Gary R. Graves; e ao Museu de História Natural de Taubaté, em nome de Herculano M. F. Alvarenga. Por fim, à FAPESP (Processo 97/05672-2) pelo apoio financeiro.
REFERÊNCIAS
BAUMEL, J.; WITMER L. M. Osteologia. In; BAUMEL, J.; KING, A. S.; BREAZILE, J. E.; EVANS, H. E; BERGE, J. C. van den. (Eds.). Handbook of avian anatomy, nomina anatomica avium. 2. ed. Cambnge: Nuttall Ornithological Club, 1993. 779p.
BAUMEL, J.; KING, A. S.; BREAZILE, J. E.; EVANS, H. E; BERGE, J. C van den. Handbook of avian anatomy: nomina anatomica avium. 2. ed. Cambrige: Nuttall Ornithological Club, 1993. 779p.
BEDDARD, F. E. On the structural characteres and classification of the cuckoos. Proc. Zool. Soc. Lond., London, 1885, p. 168-187.
BEDDARD, F. E. On the anatomy of the radiated fruit-Cuckoo (Carpococcyx radiatus). Ibis, v. 8, n. 1, p. 200-214, 1901.
BERGER, A. J. Some anatomical characteres of the Cuculidae and the Musophagidae. Wilson Bull., v. 72, p. 60-140, 1960.
BOCK, W. J. Secundary articulation of the avian mandible. Auk. Washington, v. 97. n. 1, p. 19-55, 1960.
BOCK, W. J. Kinetics of the avian skull. J. Morphol., New York, v. 114, n. 1, p, 1-52, 1964.
BRUSH, A. H. & WITT, H. H. Intraordinal relationships of the Pelecamformes and Cuculiformes: electrophoresis of feather keratins. Ibis, London, v.125, n. 1, p. 181-199, 1983.
BURTON, P. J. K. Some observations of the os unánatum in the Musophagidae. Ostrich, n. 8, p. 7-13, 1970
CRACRAFT, J. The lacrimal-ectethmoid bone complex in birds: a single character analysis. Am. Midi. Nat., New York, v. 80, n. 2, p. 316-359, 1968.
De PINNA, M. C. C. Ontogeny, rooting and polarity. In: Scotland, R. W.; Siebert, D. J. & William, D. M. (eds.). Models in phylogeny reconstruction. Londres: Clarendon Press, Oxford, 1994,p. 157-172.
DONATELLI, R. J. Cranial osteology and myology of the jaw apparatus in the Galbulidae (Aves, Piciformes). Arq. Zool., São Paulo, v. 32, n. 1, p. 1-32, 1992.
DONATELLI, R. J. The jaw apparatus of the Neotropical and of the Afrotropical woodpeckers (Aves: Piciformes). Arq. Zool., São Paulo, v. 33, n. 1, p. 1-70, 1996.
DONATELLI, R. J. Osteolog ía e miologia cranianas de Dendrocolaptidae (Passeriformes, Tyranni) 1. Géneros Glyphorynchus, Campylorhamphus, Dendrocinda, Xiphorhynchus e Dendrocolaptes. Ararajuba, Rio de Janeiro, v. 5. n. 1, p. 19-37, 1997.
FEDUCCIA, A. Evolutionary trends in the neotropical ovenbirds and woodhewers. Ornithol. Monogr., Washmgton, n. 13, p. 1-69, 1973.
FEDUCCIA, A. The Origin and Evolution of Birds. New Haven e London: Yale University Press, 1996. 420p.
FÜRBRINGER, M. Untersuchumgen zur morphologic und Systematik der Vögel. Amsterdan: Von Holkema, 1888. 1751p.
GADOW, H. Classification of birds. Proc. Zool. Soc. Lond., London, 248p. 1892.
GARROD, A. H. On the value in classification of a pecualianty in the anterior margin of the nasal bones of certain birds. Proc. Zool. Soc. Lond., London, v. 30, n. 3, p. 33-38, 1873.
HÖFLING, E. Anatomia do crânio e da cintura escapular dos Ramphastidae (Aves: Piciformes) e de alguns grupos próximos, com implicações sistemáticas. Tese (Livre-Docência). Universidade de São Paulo, 1995.
HÖFLING, E.; GASC, J. P. Biomécanique du crâne et du bec chez Ramphastos (Aves, Ramphastidae). Gegenbaurs Morphol. Jahrb., Leipzig, v. 130, n. 1, p. 125-147, 1984.
HUGHES, J. M. Phylogenetic analysis of the Cuculidae (Aves, Cuculiformes) using behavioral and ecological characteres. Auk, Washington, v. 113, n. 1, p. 10-22, 1996.
HUGHES, J. M. Monophyly and phylogeny of cuckoos (Aves, Cuculidae) inferred from osteological characters. Zool. J. Lin. Soc, n. 130, p. 263-307, 2000.
HUXLEY, T. H. On the classification of birds. Proc. Zool. Soc. Lond., London, v. 27, n. 27, p. 415-472, 1867.
HUXLEY, T. H. On the classification and distribuition of the Alectoromorphae and Heteromorphae. Proc. Zool. Soc. Lond.. London, n. 28, p. 294-319, 1868.
JOLLIE, M. The head skeleton of the chickens and remarks on the anatomy of this region in other birds. J. Morphol., New York, v. 100, n. 4, p. 389-436, 1957.
KLUGE, A. G. Ontogeny and phylogenetic systematics. Cladistics, v. 1, n. 1, p. 13-27, 1985.
KRAUS, F. An empirical evaluation of the use of the ontogeny polarization criterion in phylogenetic inference. Syst. Zool., v. 37, n. 2, p. 106-141, 1988.
LADEIRA, L. M. C. E. B. Osteologia craniana de Bucconidae (Aves: Piciformes). Dissertação (Mestrado). Universidade de São Paulo, 1997.
LOWE, P. R. On the anatomical differences between the Cuculidae and the Musophagidae. Íbis, London, n. 85, p. 490-515, 1943.
MARCELIANO, Maria Luiza V. Estudo osteol ógico e miológico do crânio de Opisthocomus hoazin (Muller, 1776) (Aves: Opisthocomidae) comparado com algumas espécies de Cracidae,Musophagidae e Cuculidae. Bol. Mus. Para. Emílio Goeldi, v. 12, n. 2, p. 95-246, 1996.
McKITRIC, M. C. Phylogenetic analysis of avian hindlimb musculature. Misc. Publ. Mus. Zool. Univ. Mich., n. 179, p. 1-87, 1991.
MÉNDEZ, A. C. Osteologia craniana de Alcedinidae Neotropicais (Aves: Coraciiformes). Dissertação (Mestrado). Universidade de São Paulo. 1998.
MORIOKA, H. Jaw musculature of swifts (Aves, Apodidae). Bull. Sci. Mus., v. 17, n. 1, p. 1-16, 1974.
NELSON, G. Ontogeny, phylogeny, paleontology and the biogenetic law. Syst. Zool., n. 27, p. 324-345, 1978.
PARKER, W. K. 1890. apud JOLLIE, M. The head skeleton of the chicken and remarks on the anatomy of this region in other birds. J. Morph., v. 100, n. 3, p. 389-436, 1957.
PAYNE, R. B. Order Cuculiformes. In: HOYO, J. del; ELLIOTT, A.; SARGATAL, J. (Eds.) Handbook of the birds of the world. Barcelona: Lynx Edictions, 1997. v. 4, p. 480-607.
PETERS, J. L. Checklist of birds of the world. Mus. Comp. Zool, Cambrige, v. 4, p. 12-76, 1940.
POSSO, S. R. Osteologia craniana dos Cuculiformes do Novo Mundo (Aves) com implicações sistemáticas. Dissertação (Mestrado). Universidade Estadual Paulista. 1999.
POSSO, S. R.; R. J. DONATELLI. Cranial osteology and systematics implications in Crotophaginae (Aves, Cuculidae). J. Zool. Syst. Evol. Research, n. 39, p. 1-10, 2001.
PYCRAFT, W. P. Contributions to the osteology of birds. Pt. VI, Cuculiformes. Proc. Zool. Soc. Lond., London, v. 73, p. 258-291, 1903.
RICHARDS, L. P.; BOCK, W. J. Functional anatomy and adaptative evolution of the feeding apparatus in the Hawaiian Honeycreeper genus Loxops (Drepanididae). Ornithol. Monogr., Washington, n. 15, p. 1-173, 1973.
SEIBEL, D. E. A phylogenetic analysis of the Cuculiformes and Opisthocomus, based on post-cranial skeletal characteres. Tese (Doutorado). University of Kansas. 1988.
SHUFELDT, R. W. The osteology of the cuckoos. Proc. Ann. Phil. Soc., n. 19, p. 4-51, 1901.
SIBLEY, G. C.; AHLQUIST, J. E. Phylogeny and classification of birds. New Haven e London: Yale University Press, 1990. 976p.
SIBLEY, G. C.; MONROE Jr., B. L. Distribution and taxonomy of birds of the world. New Haven e London: Yale University Press, 1990. 1111 p.
SICK, H. Ornitologia brasileira. 2.ed. Rio de Janeiro: Ed. Nova Fronteira, 1997. 912p
THOMPSON, D. W. On characteristic points in the cranial osteology of the parrots. Proc. Zool. Soc. Lond., London, 1899, p. 9-46.
WAKE, D. B. Phylogenetic implications of ontogenetic data. Geobios, n. 12, p. 369-378, 1989.
WHEELER, Q. D. Ontogeny and character phylogeny. Cladistics, n. 6, p. 225-268, 1990.
ZUSI, R. L. A functional and evolutionary analysis of rhynchokmesis in birds. Smithson. Contrib. Zool., n. 395, p. 1-40, 1984.
ZUSI, R. L. Patterns of diversity in the avian skull. In: HANKEN, J.: HALL, B. K. (Eds.) The skull. Chicago: Chicago University Press, 1993. v. 2, p. 391-437.
 Endereço para correspondência:
Endereço para correspondência:
Museu Paraense Emílio Goeldi
Editor do Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais
Av. Magalhães Barata, 376
São Braz – CEP 66040-170
Caixa Postal 399
Telefone/fax: 55-91- 3249 -1141
E-mail:boletim@museu-goeldi.br
Recebido: 14/04/2000
Aprovado: 17/05/2002
