










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.2 n.2 Belém ago. 2007
Florística e fitossociologia de um trecho de um hectare de floresta de terra firme, em Caracaraí, Roraima, Brasil1
Floristic composition and structure in one hectare of terra firme forest, Caracaraí, state of Roraima, Brazil
Juan Gabriel Soler AlarcónI; Ariane Luna PeixotoII
IInstituto de Pesquisas Jardim Botânico do Rio de Janeiro. Rio de Janeiro, Rio de Janeiro, Brasil (juan@jbrj.gov.br)
IIInstituto de Pesquisas Jardim Botânico do Rio de Janeiro. Rio de Janeiro, Rio de Janeiro, Brasil (ariane@jbrj.gov.br)
RESUMO
São apresentados dados florísticos e fitossociológicos de 1 ha de floresta de terra firme localizada na região do baixo rio Branco (01o01'43"S, 62o05'21"W), Roraima, Brasil. A amostragem incluiu os indivíduos arbóreos, lianas e hemiepífitas com DAP ≥ 10 cm. A altura média da floresta é de 16,63 m, com árvores emergentes que alcançaram alturas de 58, 48 e 47 m. Foram encontrados 544 indivíduos pertencentes a 194 espécies, das quais cinco estavam representadas por lianas e duas por hemiepífitas. As famílias de maior riqueza específica foram Leguminoseae, Cecropiaceae, Burseraceae, Chrysobalanaceae e Moraceae. Os gêneros de maior riqueza específica foram Pourouma, Inga, Licania, Protium e Pouteria. As espécies com densidades maiores foram Clathrotropis macrocarpa, Bocageopsis multiflora, Eschweilera coriacea, Euterpe precatoria, Inga alba, Pourouma cf. tomentosa subsp. apiculata. Dezessete famílias e 76 gêneros estiveram representados por apenas uma espécie; 40 gêneros e 104 espécies representados por apenas um indivíduo. As famílias com maiores valor de importância (VI) foram Leguminosae, Cecropiaceae, Lecythidaceae, Annonaceae e Arecaceae. Os maiores VI para espécies foram de Clathrotropis macrocarpa, Goupia glabra, Bocageopsis multiflora, Eschweilera coriacea, Euterpe precatoria. A área basal total foi 26,35 m2. O índice de Shannon (H') 4,66 e a equabilidade (J) 0,88. A região de interfúlvio entre o rio Branco e o rio Negro, em relação à flora, era dita como insuficientemente conhecida, mas de provável importância. Os dados e informações aqui apresentados e discutidos apontam a importância florística da floresta de terra firme da área.
Palavras-chave: Inventário florístico. Estrutura fitosociológica. Floresta de terra firme. Amazônia brasileira.
ABSTRACT
A quantitative floristic and phytosociological inventory in a 1 ha of terra firme forest were conducted in the region of the lower rio Branco (01o01'43"S, 62o05'21"W), Roraima, Brazil. This study included trees, lianas and hemiepiphytes with DBH ≥ 10 cm. The average height of the forest was 16,63 m, with canopy trees of 58, 48 e 47 m. The plot had 544 individuals, 194 species, of which five were represented by lianas and two by hemiepiphytes. The richest families were Leguminoseae, Cecropiaceae, Burseraceae, Chrysobalanaceae and Moraceae. The richest genera were Pourouma, Inga, Licania, Protium e Pouteria. The species with highest densities were Clathrotropis macrocarpa, Bocageopsis multiflora, Eschweilera coriacea, Euterpe precatoria, Inga alba, Pourouma cf. tomentosa subsp. apiculata. A total of 17 families and 76 genera were represented by only one individual; 40 genera and 104 species by only one individual. The families with the highest important value (IV) were Leguminosae, Cecropiaceae, Lecythidaceae, Annonaceae e Arecaceae. The highest IV for the species was found in Clathrotropis macrocarpa, Goupia glabra, Bocageopsis multiflora, Eschweilera coriacea, Euterpe precatoria. The basal area was 26,35 m2. The Shannon index (H') found was 4,66 and the equitability (J) 0,88. This region, which is localized between the rivers Branco and Negro, is said to have been poorly studied with respect of the flora but of probable importance. The data and information presented shows the floristic importance of the forest of terra firme in this area and looks forward to provide tools that will allow to compare this study with similar ones carried out in the Amazon.
Keywords: Floristic composition. Structure phytosociology. Terra firme forest. Brazilian Amazon.
INTRODUÇÃO
A região amazônica ocupa uma área aproximada de 6 milhões de km2 na América do Sul, dos quais cerca de 60% encontram-se em território brasileiro (Pires, 1980). O relevo é um dos fatores principais para determinar o tipo de vegetação, a qual está dividida em dois grupos principais: a vegetação de terra firme e a vegetação que sofre inundação (Pires; Prance, 1985).
Outra forma prática para expressar as diferenças entre tipos de vegetação é correlacionar as diferenças entre índices de biomassa: tipos de vegetação similares contém aproximadamente a mesma biomassa (área basal por hectare) (Pires; Prance, 1985). Com estes dois fatores, Pires e Prance (1985) classificam os principais tipos de vegetação da Amazônia. O presente estudo foi desenvolvido em floresta densa de terra firme, segundo esta classificação, e muitas vezes chamada na literatura simplesmente de floresta de terra firme (Milliken, 1998; Campbell et al., 1986; Ferreira; Prance, 1999, entre outros).
Embora a literatura hoje disponível sobre inventários florísticos e fitossociológicos em florestas de terra firme, várzea e igapó seja considerável, muitas vezes, é difícil comparar os resultados entre os diferentes inventários devido à diferença entre os métodos empregados, os critérios de inclusão de indivíduos e a grande diversidade local.
Campbell et al. (1986) tecem comentários sobre a dificuldade de fazer comparações entre diferentes inventários realizados na Amazônia porque muitos deles utilizam distintos critérios de inclusão. Apresentam uma tabela contendo 20 inventários realizados, incluindo o deles, que amostraram entre 0,5 e 3,8 ha com formato de área variado e critério de inclusão entre 8 e 15 cm de diâmetro a 1,3 m do solo (DAP).
Oliveira e Nelson (2001) fazem uma análise da composição florística em nível genérico entre 12 localidades da Amazônia brasileira, uma na Bolívia e uma na costa atlântica brasileira, com base em inventários publicados. Não analisaram os dados em nível específico por reconhecerem a existência de problemas de identificação botânica e nomenclatura das espécies.
Nelson e Oliveira (2001), escrevendo sobre o estado de conhecimento florístico da Amazônia, comentam sobre inventários quantitativos de árvores em florestas de terra firme e em florestas periodicamente inundadas, desde os primeiros realizados na região, em 1934, até os mais recentes. Apresentam uma tabela síntese de 37 destes inventários. Também comentam sobre as dificuldades nas comparações entre os diferentes estudos devido à variedade de metodologias, principalmente quanto à forma e tamanho da área amostral e diâmetro de inclusão. Ressaltam as dificuldades de se obter boas amostras de material testemunho e a complexidade de identificações destas amostras, o que dificulta ainda mais as comparações florísticas e estruturais ao nível de espécies.
Embora já seja significativo o conhecimento sobre a florística e a estrutura da floresta amazônica, quando se considera o tamanho e a diversidade biológica da região, a informação ainda é insuficiente para uma interpretação satisfatória da fitogeografia e da fitossociologia da região. Na revisão de literatura sobre inventários apresentada por Nelson e Oliveira, não há nenhuma citação para trabalhos realizados em Roraima. Entretanto, a região de interfúlvio rio Branco - rio Negro, onde o presente trabalho foi realizado, é apontada como área de alta importância biológica, prioritária para biodiversidade, com ausência de unidades de conservação e reconhecida como de alta importância biológica para aves, mamíferos, répteis e anfíbios. Suas florestas também são ditas como prioritárias para inventários e reconhecidas como área insuficientemente conhecida, mas de provável importância (Capobianco, 2001).
O presente estudo objetivou a busca de conhecimento sobre a estrutura e a composição florística de 1 ha da floresta de terra firme, considerando-se apenas árvores, lianas e hemiepífitas arbóreas com DAP ≥ 10 cm. A pesquisa integra um estudo etnobotânico que visou a identificar a utilização das espécies desse mesmo trecho de floresta pela comunidade ribeirinha da vila de Caicubi; e responder duas questões principais: quais as espécies que ocorrem no trecho estudado e como estão representadas suas populações (dentro dos parâmetros de inclusão estabelecidos). Os dados e informações aqui apresentados e discutidos visaram a contribuir, também, para uma futura interpretação mais acurada do vasto espaço florestado da Amazônia, fornecendo ferramentas para a conservação.
MATERIAIS E MÉTODOS
O estudo foi realizado em 1 ha de floresta de terra firme, localizado próximo à vila Caicubi (com coordenada central de 01o01'43"S e 62o05'21"W, altitude de 50 msnm). Esta vila, habitada por 400 pessoas em 72 famílias (Orange Ferreira, comunicação pessoal), localiza-se na margem do igarapé Caicubi, afluente do rio Jufari, na região do baixo rio Branco (interflúvio rio Branco - rio Negro), município de Caracaraí, Roraima (Figura 1). As cidades mais próximas à vila de Caicubi são Barcelos e Manaus, localizadas no rio Negro, e Caracaraí, em Roraima, no rio Branco. Segundo a classificação de Veloso et al. (1991), a área de estudo é coberta por floresta ombrófila densa das terras baixas e, segundo a classificação de Pires e Prance (1985), floresta densa de terra firme.
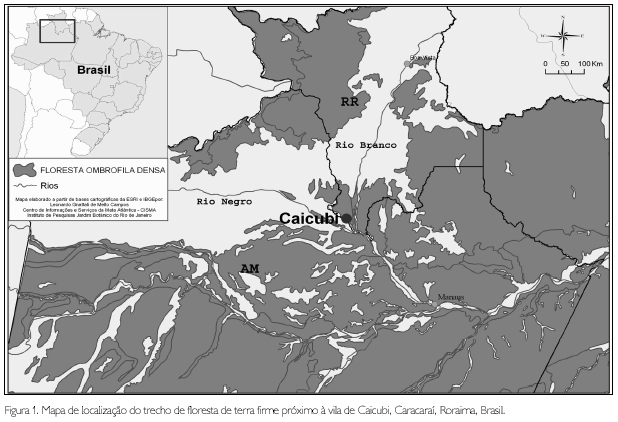
O clima da região é tipicamente quente e úmido (Affi de Köepen). Vila Nova et al. (1976) apresentam dados climáticos de Barcelos (0o59'S, 62o55'W), onde se localiza a estação meteorológica mais próxima a Caicubi, e apontam, em uma série de 30 anos (1931 a 1960), temperatura média anual de 26oC; pluviosidade média anual de 1999 mm, sendo os meses mais chuvosos abril, maio e junho; umidade relativa de 86%. O trecho estudado pertence à Formação Solimões; geomorfologicamente está classificado como interflúvio tabular de relevo de topo aplainado e solo classificado como Latossolo Amarelo Álico (RADAMBRASIL, 1978).
A escolha do trecho a ser estudado foi feita numa área que, de acordo como a população local, teve pouca intervenção, sendo apenas extraídos produtos não madeiráveis, principalmente açaí (Euterpe precatoria), bacaba (Oenocarpus bacaba), cipó-titica (Heteropsis cf. flexuosa), arumã (Ishnosiphon sp.), castanha (Bertholletia excelsa) e ubim (Geonoma spp.).
O método escolhido para o estudo florístico e fitossociológico foi o de parcelas, de modo geral, empregado em estudos similares na Amazônia (Campbell et al., 1986; Milliken, 1998; Ferreira; Prance, 1999, entre outros). Na área previamente escolhida, delimitou-se 1 ha em forma de retângulo (200 m x 50 m) em direção Norte-Sul subdividido em parcelas de 10 m x 10 m. Optou-se por um retângulo, pois ao tentar-se delimitar um quadrado de 100 m x 100 m, foram encontrados castanhais nos quais a população local coleta frutos e para tal limpam parcialmente um trecho correspondente ao diâmetro das copas das castanheiras. A diferença de forma da parcela entre quadrado e retângulo, das dimensões empregadas, parece não influenciar significativamente em análises relativas à riqueza de espécies; retângulos mais estreitos (10 m x 1000 m), entretanto, podem capturar maior riqueza (Laurance et al., 1998).
Dentro das parcelas, foram numerados, seqüencialmente, com pequenas placas de material plástico, todos os indivíduos arbóreos, lianas e hemiepífitas com DAP ≥ 10 cm. Quando as árvores apresentavam sapopemas, o DAP era medido acima destas. Para todos os indivíduos, foram anotados, em planilhas, dados de altura total, altura do fuste, DAP e características morfológicas, como cor do ritidoma, exsudatos (látex, seiva, resina, goma), odor ou outros caracteres que pudessem auxiliar na identificação, bem como o nome popular. Entre novembro de 2003 e fevereiro de 2004, foram coletadas amostras das morfoespécies para identificação e depósito em herbários. A coleta foi realizada com tesoura de alta poda e, para os indivíduos com copas acima de 14 m, foi necessário escalar as árvores utilizando peconha, técnica amplamente utilizada pelos habitantes da região, para a coleta de alguns produtos das palmeiras de açaí (Euterpe precatoria), bacaba (Oenocarpus bacaba) e patauá (Oenocarpus bataua), entre outras. As amostras botânicas foram prensadas em jornal e preservadas em álcool durante todo o período de permanência no campo; após, foram secas em estufa nos laboratórios do Instituto Nacional de Pesquisas da Amazônia (INPA). A coleção completa das amostras foi depositada no herbário do Jardim Botânico do Rio de Janeiro (RB) e duplicatas de exemplares férteis foram depositadas no herbário do INPA.
A identificação taxonómica foi realizada inicialmente no campo, utilizando-se os livros de Gentry (1993) e Ribeiro et al. (1999), muito empregados em identificação de espécies vegetais. Após esta etapa, as identificações foram confirmadas com literatura especializada e comparando-se as amostras com os exemplares depositados nos herbários do INPA e RB. Especialistas em taxonomia confirmaram ou retificaram as identificações das famílias Lauraceae, Burseraceae, Moraceae e Leguminoseae. Não foi possível identificar três espécimes que foram tratados como um grupo separado, denominado de 'indeterminadas'. Estes espécimes permaneceram desfolhados durante toda a permanência em campo, o que impossibilitou também a coleta para constituir exemplares testemunhos. Três morfoespécies idênticas a táxons ainda não descritos formalmente, porém citados em Ribeiro et al. (1999), como Ocotea sp. E, Aniba aff. Williamsii e Qualea sp.1, são aqui citadas dessa forma.
Seguiu-se o sistema de classificação de Cronquist (1981), excetuando Leguminosae, que foi considerada como família única, como indicado por Polhill e Raven (1981). Optou-se por este procedimento para possibilitar uma melhor comparação entre o trecho estudado e outros estudos realizados na região amazônica. A grafia dos nomes das espécies foi conferida em revisões taxonómicas ou no Index Kewensis, versão on line. Os nomes dos autores das espécies encontram-se abreviados segundo Brummit e Powel (1992). Os nomes populares foram indicados por 11 colaboradores da vila de Caicubi que auxiliaram nos trabalhos de campo.
O programa Fitopac 1 (Shepherd, 1995) foi utilizado para analisar os parâmetros fitossociológicos. No caso das espécies e das famílias, foram calculados os valores absolutos e relativos de densidade, freqüência e dominância e os valores de importância (VI) (Müller-Dombois; Ellenberg, 1974), assim como o índice de diversidade de Shannon (H') e a equabilidade (J) (Magurran, 1988).
RESULTADOS E DISCUSSÃO
No trecho de 1 ha de floresta de terra firme, foram encontrados 544 indivíduos, distribuídos em 43 famílias, 106 gêneros e 192 espécies. Em realção ao hábito, 185 eram de espécies arbóreas (em 100 gêneros), seguido pelas lianas com cinco espécies (em quatro gêneros e uma indeterminada) e hemiepífitas com duas espécies (em dois gêneros). Na Tabela 1, são apresentadas as espécies, em ordem alfabética de famílias, seus nomes populares e o número de coleta do exemplar testemunho.
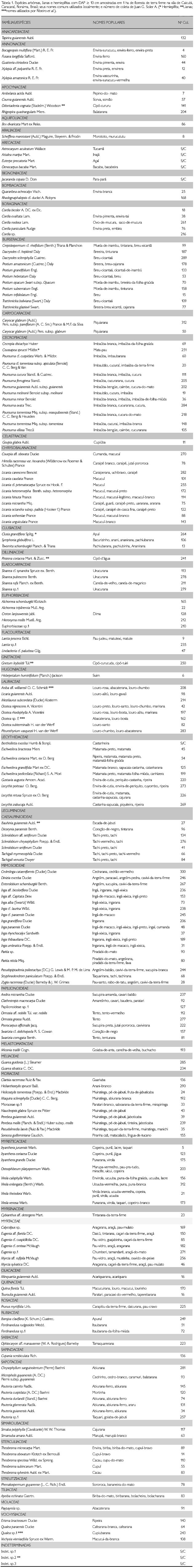
A família com maior riqueza específica foi Leguminosae, com 32 espécies, 17 delas pertencentes à subfamília Mimosoideae, oito à subfamília Papilionoideae e sete à subfamília Caesalpinoideae. As outras famílias com maior riqueza de espécies foram Cecropiaceae (13 espécies), Burseraceae (11), Chrysobalanaceae (11) e Moraceae (10). Foram encontradas 17 famílias representadas por apenas uma espécie.
A família Leguminosae também se caracterizou por apresentar o maior número de indivíduos (102), pertencendo 46 deles à subfamília Papilionoideae, 43 à Mimosoideae e 13 à Caesalpinioideae. As outras famílias com maior número de indivíduos foram Cecropiaceae (51), Lecythidaceae (46), Annonaceae (42) e Arecaceae (32). Foram encontradas dez famílias representadas por apenas um indivíduo (Tabela 2).
Os gêneros que apresentaram maior riqueza de espécies foram Pourouma (11), Inga (dez), Licania (nove), Protium (sete) e Pouteria (seis). Estiveram representados por apenas uma espécie 76 gêneros. Em relação ao número de indivíduos por gênero, Pourouma foi o mais abundante, com 45 indivíduos, seguido por Clathrotropis (39), Eschweilera (35), Bocageopsis (30) e Inga (29). Gêneros representados por apenas um indivíduo totalizaram 40.
O gênero Pourouma destacou-se como mais abundante porque somou os indivíduos de dois táxons bem representados na área, P cf. tomentosa subsp. apiculata (15 indivíduos) e P. minor (13). As outras nove espécies estiveram assim representadas: duas espécies com quatro indivíduos, duas espécies com dois indivíduos e cinco espécies com apenas um indivíduo.
As espécies com maior número de indivíduos foram Clathrotropis macrocarpa (39), Bocageopsis multiflora (30), Eschweilera coriacea (21), Euterpe precatoria (20), Inga alba (15) e Pourouma cf. tomentosa subsp. apiculata (15). Estas seis espécies representaram 3% do total de espécies, porém, 25,7% dos indivíduos amostrados (Figura 2).
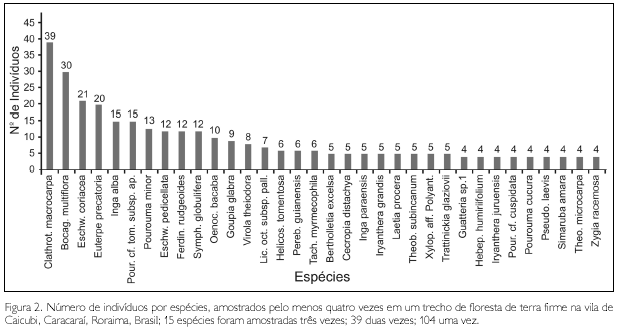
Do total de 192 espécies, apenas 11 estiveram representadas por dez ou mais indivíduos, as quais, juntas, representaram 38,2% dos indivíduos. Também é interessante ressaltar que as 25 espécies com densidades maiores representam 51,6% do total dos indivíduos. Isto corrobora com Pires e Prance (1985), quando afirmam que não existe uma espécie dominante nas florestas úmidas tropicais; entretanto, sempre vai existir um grupo de espécies dominantes, entre cinco a dez ou, às vezes, até 30, que quando somados seus indivíduos ultrapassam 50% do total.
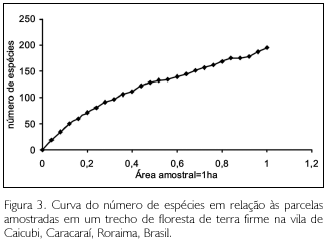
O surgimento progressivo de novas espécies amostradas encontra-se representado na curva espécies/área (curva do coletor) da Figura 3. O número de espécies amostradas em cada subparcela de 10 m x 10 m variou de 1 a 10. Observase que, embora tenha havido dois pontos de aparente estabilidade entre 0,52 a 0,56 ha e entre 0,85 a 0,88 ha, não há, no final da amostragem, uma estabilidade, demonstrando que a área é insuficiente para uma boa representação da riqueza da floresta de terra firme. Tal informação corrobora com pesquisas realizadas por Campbell etal. (1986) e Milliken (1998), que observaram também ser insuficiente os dados de 1 ha na floresta de terra firme da Amazônia para uma avaliação acurada da riqueza de espécies.
As espécies representadas por apenas um indivíduo somam 104, ou seja, 54,2% do total (Figura 4). As cinco espécies de lianas e as duas espécies de hemiepífitas amostradas estiveram representadas por apenas um indivíduo. Considerando como espécies raras aquelas representadas por apenas um indivíduo em 1 ha, Martins (1991) compara alguns trabalhos realizados em florestas brasileiras e cita três inventários em áreas de 1 ha na Amazônia que utilizaram, como critério de inclusão, árvores com DAP ≥ 10 cm: Pires et al. (1953), em floresta de terra firme em Castanhal, Pará, encontraram 45 espécies raras (25,14%); Black et al. (1950), em floresta de terra firme em Tefé, Amazonas, encontraram 42 de espécies raras (53,16%); e Porto et al. (1976), em 'mata de baixio' em Manaus, Amazonas, encontraram 58 espécies raras (50,88%).

O índice de Shannon (H') encontrado foi 4,652 e a equitabilidade (J) 0,885. Os altos valores para o índice de Shannon e de equitabilidade podem ser atribuídos ao grande número de espécies raras (54,2%). Considerando apenas as espécies arbóreas, os valores para H' e J foram 4,616 e 0,884, respectivamente. O número de espécies raras encontrado em Caicubi, maior do que qualquer outro citado anteriormente, e os altos valores do índice de diversidade e da equitabilidade corroboram com o comentário exposto em Capobianco (2001), que o interflúvio entre os rios Branco e Negro é uma área de alta diversidade e estudos são necessários para inventariar a sua biota.
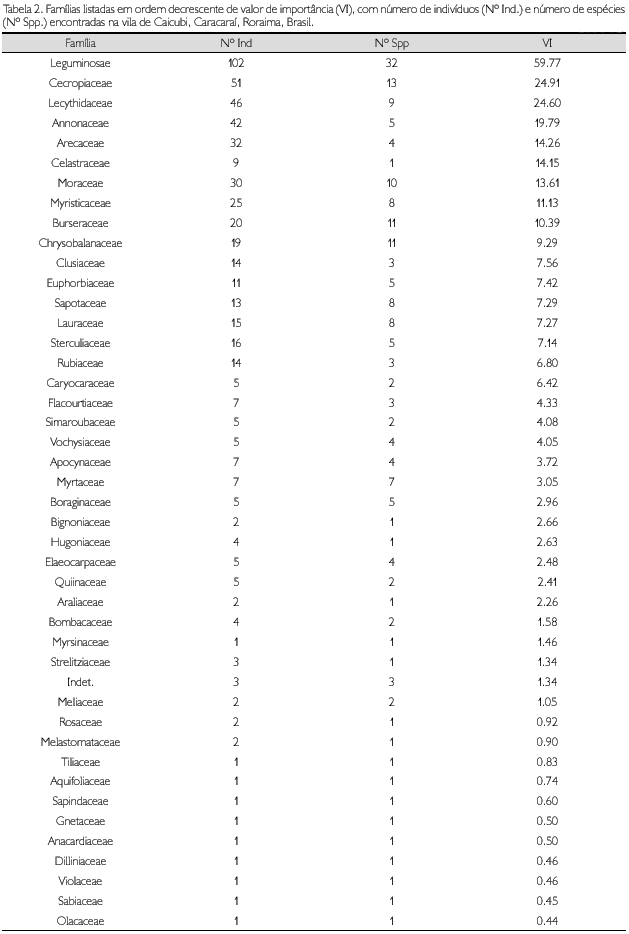
As cinco famílias de mais alto VI no trecho estudado foram Leguminosae (59,77), Cecropiaceae (24,91), Lecythidaceae (24,60), Annonaceae (19,79) e Arecaceae (14,26) (Tabela 2).
Na Tabela 3 são apresentadas as espécies com seus respectivos parâmetros fitossociológicos, em ordem decrescente do VI. As cinco espécies com maior VI foram Clathrotropis macrocarpa, Goupia glabra, Bocageopsis multiflora, Eschweilera coriacea e Euterpe precatoria, representando, juntas, 21,87% dos indivíduos amostrados e 23,8% da área basal. C. macrocarpa foi a espécie de maior VI pela sua abundância (39 indivíduos), tendo alcançado as maiores densidade e freqüência relativas. C. macrocarpa, árvore de porte médio, teve a variação na altura total de 8 a 28 m e sua altura média foi de 16 m; o DAP variou de 10,2 a 37,9 cm e DAP médio de 20 cm. Bocageopsis multiflora, Eschweilera coriacea e Euterpe precatoria, com 30, 21 e 20 indivíduos, respectivamente, também têm destaque por sua abundância e freqüência na área estudada. Entretanto, Goupia glabra, com nove indivíduos, e Cedrelinga cataeniformis, com apenas um indivíduo, têm valores baixos de densidade e freqüência relativas, apresentando os maiores valores de dominância relativa (DoR), 10,44% e 7,55%, respectivamente, o que as coloca entre as seis espécies de maiores VI. Este padrão também se repete para Caryocarglabrum subsp. parvifolium, com três indivíduos e DoR 4,26%; Parkia nitida com três indivíduos e DoR 3,26%; e Ormosia grossa com um indivíduo e DoR 3,98%.
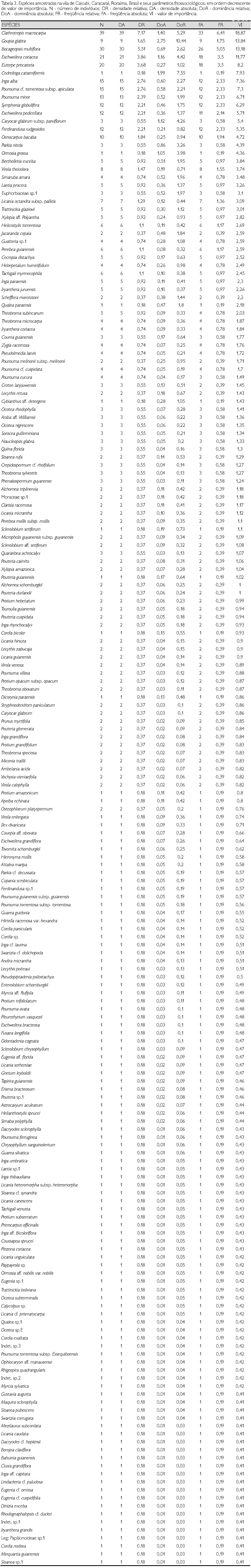
Além das quatro espécies de maiores populações (C. macrocarpa, B. multiflora, E. coriaceae e E. precatoria), as espécies Inga alba (15 indivíduos), Pourouma cf. tomentonsa supbsp. apiculata (15), Pourouma minor (13), Eschweilera pedicellata (12), Symphonia globulifera (12), Ferdinandusa rudgeoides (12) e Oenocarpus bacaba (10) apresentaram populações com mais de dez indivíduos e valores elevados na freqüência relativa.
Os 544 indivíduos amostrados, dentro dos parâmetros estabelecidos, somaram uma área basal de 26,35 m2. Pires e Prance (1985) citam que florestas densas podem ultrapassar 40 m2 de área basal e que florestas abertas encontram-se entre 18 a 24 m2. Os resultados indicam que se trata de uma floresta densa com uma baixa área basal aproximando-se de florestas abertas.
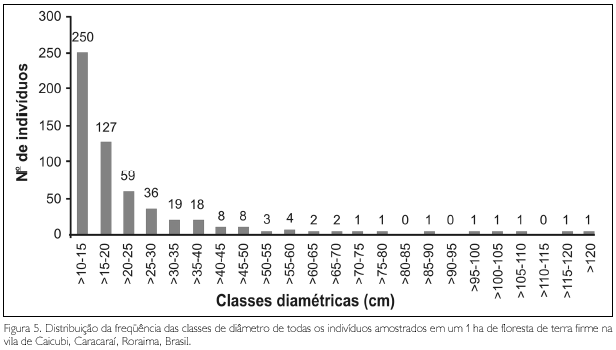
Na distribuição das árvores por classes de diâmetro (Figura 5), observa-se uma predominância de indivíduos (45,96%) na classe diamétrica de 10-15 cm; 23,35% dos indivíduos estão inseridos na classe de >15-20 cm; 10,85% na classe >20-25 cm; 6,6% na classe de >25-30; 3,49% na classe de >30-35; 3,3% na classe >35-40; os demais, totalizando 6,43%, dividem-se em classes diamétricas acima de 40 cm. O maior diâmetro foi de 159,15 cm, encontrado no único indivíduo de Cedrelinga cataeniformis. O diâmetro médio foi 20,15 cm, incluindo todos os indivíduos da amostragem; excluindo as lianas, o diâmetro foi 20,24 cm. A curva formada pelos valores no conjunto de classes estabelecidas mostra a configuração de J invertido, demonstrando que há uma maior quantidade de indivíduos de menor porte e poucos indivíduos emergentes. Isto é esperado que ocorra com o incremento do diâmetro dos indivíduos, pois a distribuição de classes de tamanho é conseqüência da dinâmica da floresta, onde a quantidade de espaço constringe o número de árvores que podem se acomodar em um determinado tamanho de classe (Swaine, 1989).
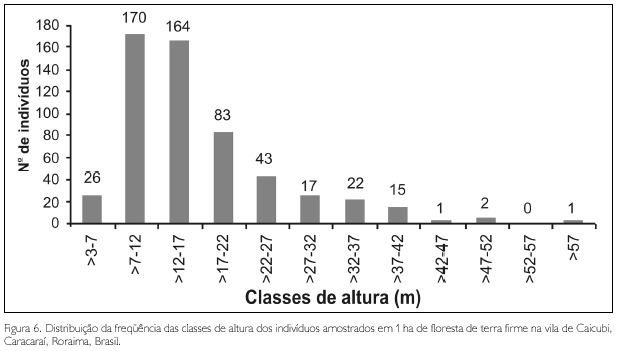
Agruparam-se os indivíduos amostrados em 12 classes de altura (Figura 6), variando a amplitude de alturas entre 3 a 58 m. O maior número de indivíduos concentrou-se na segunda (31,25%) e terceira classes (30,15%). A classe de maior altura apresentou apenas um indivíduo de Ormosia grossa, representada na área por esse único indivíduo. O pequeno número de amostras na primeira classe deveu-se ao fato de várias árvores desta classe apresentarem a copa quebrada (o que diminuiu a sua altura total), e 46% dos indivíduos desta classe possuírem alturas de 7 m, encontrando-se próximo à altura da segunda classe (>7-12 m). A altura média encontrada foi de 16,41 m (excluindo as lianas), sendo as árvores emergentes, indivíduos de Ormosia grossa (58 m de altura), Jacaranda copaia (48 m), Laetia procera (48 m) e Cedrelinga cataeniformis (47 m). J. copaia e L. procera estavam representadas por dois e cinco indivíduos, respectivamente, e as duas outras espécies por apenas um indivíduo.
Na Vila de Maré (61o15W, 1o45"S), em terras Waimin Atroari, Milliken (1998) inventariou 1 ha de floresta de terra firme, utilizando método de parcela e incluindo indivíduos com DAP ≥ 10 cm, sendo esta a área mais próxima àquela estudada em Caicubi. Encontrou 662 indivíduos, em 216 espécies e 41 famílias das quais 17 lianas em 14 espécies e oito famílias. O número de espécies e de famílias foi aproximado ao encontrado no presente estudo. O número de indivíduos, porém, foi maior (Tabela 4).
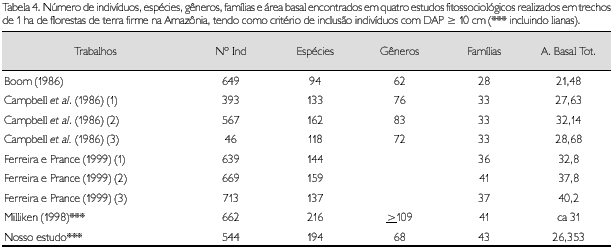
Ao comparar as dez famílias com maior VI apresentadas por Milliken (1998) com àquelas do presente estudo, sete delas são comuns, sendo Leguminosae a de maior VI em ambas as áreas. As demais aparecem em diferentes ordenações. Em relação às espécies, Cathrotropis macrocarpa é a de maior VI nas duas áreas: em Vila de Maré apresentou VI de 24,7 e 72 indivíduos; em Caicubi apresentou VI de 18,87 e 39 indivíduos. Eschweilera coriacea é a segunda espécie de maior VI em Vila de Maré (VI de 18,2 e 53 indivíduos) e é a quarta de maior VI em Caicubi (VI de 11,77 e 21 indivíduos). Além destas duas espécies, não há outras comuns entre as dez espécies de maior VI, em ambas as áreas. Dentre as dez de maior VI da área de Vila de Maré, quatro não foram amostradas em Caicubi. Na Tabela 4 estão sintetizados os dados de Vila de Maré e de outros dois estudos realizados na Amazônia Central e Sudeste.
Ferreira e Prance (1999) amostraram 3 ha de floresta alta no Parque Nacional Jaú (1o 90'-3o 00'S, 61o 25'-63o50'W), usando DAP ≥ 10 cm. Encontraram uma média de 146,7 espécies/hectare e densidade média de 673,7 árvores/hectare; o mais rico apresentou 159 espécies e o de maior densidade apresentou 713 indivíduos. Em dois dos hectares por eles amostrados, cinco das famílias de maiores VI estão incluídas entre as dez de maiores VI em Caicubi (Leguminosae por eles tratada em três famílias). No outro hectare, são seis famílias que estão entre as dez de maiores VI em Caicubi. Escweilera coriacea é a única espécie comum entre as dez de maiores VI entre um dos hectares estudados por Ferreira e Prance (1999) e em Caicubi. Esta também é a única espécie comum entre as dez de maiores VI entre o estudo de Milliken (1998) e de Ferreira e Prance (1999). Os outros dois hectares, de Ferreira e Prance (1999), não possuem espécies em comum com Caicubi, quando se observam as dez espécies de maiores VI (Tabelas 5 e 6).
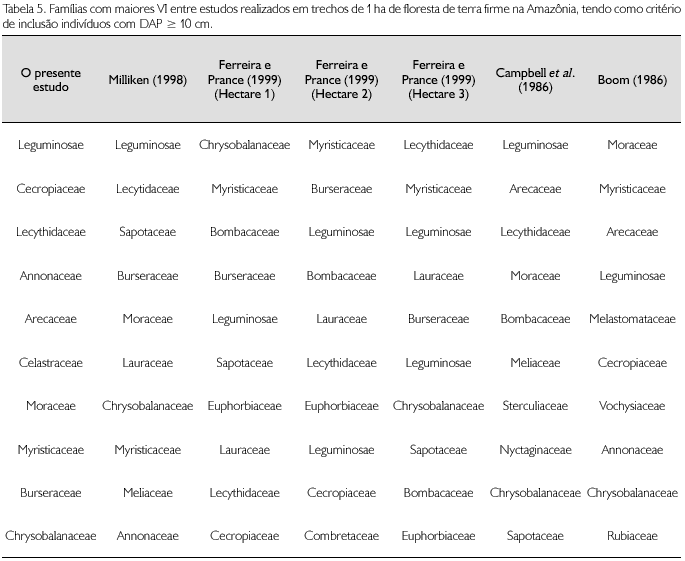
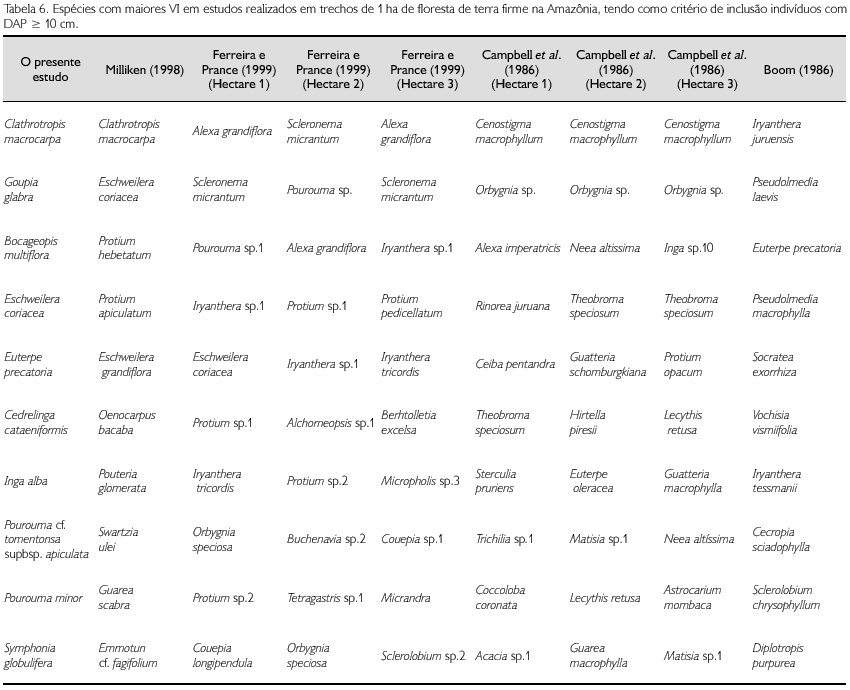
Campbell et al. (1986), em 'O Deserto', no rio Xingu, Pará (3o 29' S/ 51o 40' W), amostraram 3 ha de floresta de terra firme, utilizando também como critério de inclusão DAP ≥ 10 cm. Encontraram 393, 567 e 460 indivíduos em cada um dos hectares estudados; 133, 162 e 118 espécies, respectivamente; e 33 famílias nos três hectares estudados. Das dez famílias com maiores VI, cinco estão entre as dez de maiores VI em Caicubi. Nenhuma espécie, entre as 10 de maiores VI, é comum ao trecho estudado em Caicubi (Tabelas 5 e 6).
Boom (1986), em Beni, Bolívia (11o45'S, 66o02'W), amostrou 1 ha de floresta de terra firme, utilizando um DAP ≥ 10 cm. Encontrou 649 indivíudos, 94 espécies e 28 famílias. Das dez famílias com maiores VI, sete estão entre as dez de maiores VI em Caicubi, mas não há espécie em comum entre as dez de maiores VI (Tabelas 5 e 6).
Os dados encontrados em Caicubi e nos quatro últimos trabalhos citados corroboram com Nelson e Oliveira (2001), quando afirmam que a composição das florestas amazônicas varia em função de sua fisionomia e da distância entre sítios amostrados, e trazem aporte de dados para a confirmação da hipótese de que a floresta de terra firme é um mosaico de florestas com variada composição e estrutura.
A família Leguminosae, quando considerada como um só táxon, é, de modo geral, a de mais destaque em importância fitossociológica nos estudos citados, excetuando Boom (1986), que encontrou Moraceae como a família dominante. A família Leguminosae também é apontada como de grande importância em outros estudos realizados na Amazônia (Nelson; Oliveira, 2001). Entretanto, em relação a espécies, não há nenhuma que possa ser considerada como de destaque no conjunto de trechos estudados, o que corrobora com Campbel et al. (1986) quando afirmam que a Amazônia é um grande mosaico de florestas, e com Campbel et al. (1986) e Nelson e Oliveira (2001), quando alertam para ter cuidado ao se tentar fazer generalizações para a Amazônia a partir de dados florísticos e estruturais oriundos de amostras pontuais.
A família Lecythidaceae alcança sua maior expressão nas florestas de terra firme da Amazônia e a presença de muitas espécies dessa família pode ser considerada como indicadora de florestas preservadas (Mori, 2001). Os dados encontrados em Caicubi comprovam este fato. Foram encontradas nove espécies de árvores desta família, uma delas representadas por 21 indivíduos (Eschweilera coriacea), uma por 12 indivíduos (E. pedicellata), uma com cinco indivíduos (Bertholetia excelsa), duas com dois e quatro por apenas um indivíduo. E. coriacea é uma espécie comum e de distribuição ampla na Amazônia e apontada entre as espécies com destaque em diversos inventários (Salomão et al., 1988; Ferreira; Prance, 1999; Milliken, 1998), sendo a quarta espécie em VI no trecho estudado.
Em Caicubi, Cecropiaceae foi a segunda família de maior VI. Das 13 espécies amostradas, 11 pertencem ao gênero Pourouma, uma a Coussapoua e uma espécie de Cecropia. Cecropiaceae não aparece como família de destaque no trecho estudado por Milliken (1998), embora seja este o trecho mais próximo de Caicubi. O mesmo ocorre nos trechos estudados por Campbel et al. (1986). Em Ferreira e Prance (1999) não ocorre com destaque em um dos trechos e é a décima em VI nos outros dois trechos estudados por estes autores.
CONCLUSÃO
O trecho estudado na vila de Caicubi, Caracarí, Roraima, em termos de estrutura, diversidade e composição florística, dentro dos limites estabelecidos pela metodologia utilizada, corrobora com as características estruturais apontadas por diferentes autores para as florestas de terra firme da Amazônia. As famílias mais importantes no estudo realizado são, de modo geral, aquelas encontradas em outros trabalhos realizados na Amazônia, sendo elas Leguminosae, Cecropiaceae, Lecythidaceae, Annonaceae, Arecaceae, Moraceae, Myristicaceae, Burseraceae, Chrysobalanaceae, Sapotaceae e Lauraceae. A importância particular das Leguminosas, encontradas no trecho estudado, parece ser uma característica de florestas de terra firme da Amazônia, especialmente daquelas na bacia do rio Negro.
Entretanto, a composição de espécies encontradas difere substancialmente de outros inventários conduzidos na Amazônia, corroborando com as premissas de que a floresta de terra firme é um mosaico de florestas. Assim, embora os inventários em áreas de 1 ha, no contexto da Amazônia, não pareçam adequados para mostrar toda a diversidade de espécies, como apontado pela curva espécie/área, estudos em áreas deste tamanho podem espelhar a riqueza de espécies e as diferenças em composição de espécies de distintos trechos de floresta de terra firme.
Devido a diferenças em metodologias aplicadas em inventários na floresta de terra firme da Amazônia, é difícil fazer comparações entre resultados encontrados por diferentes autores que estudaram estas florestas. Listagens completas de espécies, seus exemplares de referência e tabelas contendo dados de estrutura são essenciais para a comparação entre os trabalhos, possibilitando conclusões mais abrangentes sobre estas florestas.
AGRADECIMENTOS
Ao instituto Caiuá, por financiar o projeto, e principalmente a Walo Leuzinger, por ter acreditado em nosso trabalho. À comunidade Caicubi, pela acolhida carinhosa, em especial a Ernane Fontes Barbosa, pela ajuda e dedicação ao campo. A C.A. Cid Ferreira, pelo inestimável apoio no herbário do INPA, bem como a Paulo Assunção, pelo auxílio na confirmação das identificações neste mesmo herbário. Aos especialistas Alberto Vicentini, Douglas Daly, José Eduardo Ribeiro, Michael Hopkins pela identificação de espécies de famílias de suas especialidades. A Rejan Guedes Bruni, pela leitura crítica do manuscrito. À CAPES, pela bolsa de mestrado concedida. Ao Programa de Pós-graduação do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, pela oportunidade de realização do trabalho.
REFERÊNCIAS
BOOM, B. M. A. Forest inventory in Amazonian Bolivia. Biotropica, v. 18, n. 4, p. 287-294, 1986.
BRUMMITT, R. K.; POWEL, C. E. Authors of plants names. Kew, UK.: Royal Botanic Gardens, 1992. 732 p.
CAMPBELL, D. G.; DALY, D. C., PRANCE, G. T.; MACIEL, U. N. Quantitative ecological inventory of terra firme and várzea tropical forest on the Rio Xingu, Brazilian Amazon. Brittonia, v. 38, n. 4. p. 369-393, 1986.
CAPOBIANCO, J. P. R. (Org.). Biodiversidade na Amazônia Brasileira. São Paulo: Instituto Socioambiental, 2001. 540 p.
CRONQUIST, A. An integrated system of classification of flowering plants: The New York Botanical Garden. New York: Columbia University Press, 1981. 1262 p.
FERREIRA, L. V.; G. T. PRANCE. Ecosystem recovery in terra firme forests after cutting and burning: a comparison on species richness, floristic composition and forest structure in the Jaú National Park, Amazonia. Botanical Jounal of the Linnean Society, v. 130, p. 97-110, 1999.
GENTRY, H. A. A Field guide to the families and genera of wood plants of Northwest South America (Colombia, Ecuador, Peru) with supplementary notes on herbaceous taxa. Washington: Conservation International, 1993. 895 p.
LAURANCE, W. F.; FERREIRA, L. V.; RANKIN-DE-MERONA, J. M.; HUTCHINGS, R. W. Influence of Plot Shape on Estimates of Tree Diversity and Community Composition in Central Amazônia. Biotropica, v. 30, n. 4, p. 662-665, 1998.
MARTINS, F. R. Estrutura de uma floresta mesófila. Campinas: UNICAMP, 1991. 246 p.
MAGURRAN, A. E. Ecological diversity and its measurement. Princeton, New Jersey: Princeton University Press, 1988. 179 p.
MILLIKEN, W. Structure and composition of one hectare of central Amazonian terra firme forest. Biotropica, v. 30, n. 4, p. 530-537, 1998.
MORI, S. A. A família da castanha-do-pará: símbolo do Rio Negro. In: OLIVEIRA, A. A.; DALY, D. C. (Eds.). Florestas do Rio Negro. [S.l.]: Companhia das Letras/UNIP 2001, p 119-141.
MUELLER-DOMBOIS, D.; ELLENBERG, H. Aims and methods of vegetation ecology. New York: John Wiley, 1974. 547 p.
NELSON, B. W.; OLIVEIRA, A. Área de Botânica. In: CAPOBIANCO, J. P. R. (Ed.) Biodiversidade na Amazônia Brasileira. São Paulo: Instituto Sócio-ambiental, 2001. p. 132-153.
OLIVEIRA, A.; NELSON, B. W. Floristic relationships of terra firme forests in the Brazilian Amazon. Forest Ecology and Management, v. 146, p. 169-179, 2001.
PIRES, J. M. Tipos de vegetação da Amazônia. Vegetalia-Escritos e Documentos, (IBILCE. UNESP) v. 4, p. 1-31, 1980.
PIRES, J. M.; PRANCE, G. T. The Amazon Forest: A Natural Heritage to be Preserved. In: PRANCE, G. T.; ELLIAS, T. S. (Orgs.). Extintion is forever. New York: Botanical Garden, 1977. p. 158-194.
PIRES, J. M.; PRANCE, G. T. The vegetation types of the Brazilian Amazon. In: Prance, G. T.; Lovejoy, T. E. (Eds.). Key environments Amazônia. New York: Pergamon Press, 1985. p. 109-145
POLHIL, R. M.; RAVEN, P. H. Advances in Legume Systematics, Part 1. Kew: Royal Botanical Garden, 1981. 425 p.
RADAMBRASIL. (Projeto Radambrasil, Ministério das Minas e Energia). Levantamento dos Recursos Naturais, Volume 18. Folha AS. 20 Manaus; Geologia, geomorfologia, pedologia, vegetação e uso potencial da terra. Rio de Janeiro, 1978. 628 p., 7 mapas.
RIBEIRO, J. E. L. S.; HOPKINS, M. J. G.; VICENTINI, A.; SOTHERS, C. A.; COSTA, M. A. S.; BRITO, J. M.; SOUZA, M. A. D.; MARTINS, L. H. P.; LOHMANN, L. G.; ASSUNÇÃO, P. A. C. L.; PEREIRA, E. C.; SILVA, C. F.; MESQUITA, M. R.; PROCÓPIO, L. C. Flora da Reserva Ducke: guia de identificação das plantas vasculares de uma floresta de terra-firme na Amazônia Central. Manaus: INPA/DFID, 1999. 800 p.
SALOMÃO, R. P.; SILVA, M. F. F.; ROSA, N. A. Inventário ecológico em floresta tropical de terra firme, Serra Norte, Carajás, Pará. Bol. Mus. Para. Emílio Goeldi, Belém, v. 4, n. 1, p. 1-46, 1988.
SHEPHERD, G. I. Fitopac 1: manual do usuário. Campinas: Universidade Estadual de Campinas, 1995. 93 p.
SWAINE, M. D. Population dynamics of tree species in tropical forests. In: HOLM-NIELSEN, L. B., NIELSEN, I. C.; BALSLEV, H. Tropical forests, botanical dynamics, speciation and diversity. Scotland: University of Aberdeen, 1989. p. 101-110.
VELOSO, H. P., RANGEL FILHO, A. L. R.; LIMA, J. C. A. Classificação da vegetação brasileira, adaptada a um sistema universal. Rio de Janeiro: Fundação IBGE, 1991. 123 p.
VILLA-NOVA, N. A.; SALATI, E.; MATSUI, E. Estimativa da evaporatranspiração na Bacia Amazônica. Acta Amazonica, Manaus, v. 6, n. 2, p. 215-228, 1976.
 Endereço para correspondência:
Endereço para correspondência:
Museu Paraense Emílio Goeldi
Editor do Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais
Av. Magalhães Barata, 376
São Braz - CEP 66040-170
Belém - PA - Brasil
Caixa Postal 399
Telefone/fax: 55-91- 3249 -1141
E-mail: boletim@museu-goeldi.br
Recebido: 09/11/2005
Aprovado: 16/03/2007
1Parte da dissertação de mestrado do primeiro autor, desenvolvida na Escola Nacional de Botânica Tropical do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, com o apoio financeiro e logístico da Fundação Caiuá de Gestão Ambiental.
