










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.2 n.3 Belém dez. 2007
Osteologia craniana de Geotrygon montana (Linnaeus, 1758) (Columbiformes: Columbidae) comparada com os Columbiformes do Novo Mundo
Cranial osteology of Geotrygon montana (Linnaeus, 1758) (Columbiformes: Columbidae) compared to other New World Columbiformes
Maria Luiza Videira MarceliandI; Reginaldo José DonatelliII; Sérgio Roberto PossoIII
IMuseu Paraense Emílio Goeldi. Coordenação de Zoologia. Belém, Pará, Brasil (mlvideira@museu-goeldi.br)
IIUniversidade Estadual Paulista. Laboratório de Vertebrados. Departamento de Ciências Biológicas. Bauru, São Paulo, Brasil (rjdonat@fc.unesp.br)
IIIUniversidade Federal de Mato Grosso do Sul. Laboratório de Ecologia, Filogenia e Conservação de Aves Neotropicais. Departamento de Ciências Naturais. Três Lagoas, Mato Grosso do Sul, Brasil (srposso@hotmail.com)
RESUMO
Estudos sobre a osteologia craniana dos Columbiformes são escassos na literatura. No que se refere a G. montana, não foram encontrados trabalhos desta natureza, sendo esta, provavelmente, a primeira descrição detalhada da anatomia craniana da espécie. Procurou-se, neste estudo, descrever a osteologia e a musculatura que movimenta as maxilas em G. montana e, com base nesta descrição, comparar com outras espécies de Columbiformes do Novo Mundo e ampliar o conhecimento anatômico nesta ordem, bem como identificar caracteres anatômicos para uma futura análise cladística. Estudou-se a osteologia craniana em 35 espécimes pertencentes a seis gêneros de Columbidae, abrangendo cerca de 70% dos gêneros de Columbiformes do Novo Mundo. Os resultados mostraram inúmeras diferenças relacionadas ao grau de desenvolvimento das estruturas entre as espécies analisadas, bem como algumas características peculiares a alguns táxons. Isso indica que a osteologia craniana dos Columbidae pode ser uma ferramenta das mais importantes para um estudo sistemático a posteriori.
Palavras-chave: Geotrygon montana. Columbidae. Crânio. Osteologia.
ABSTRACT
Studies on cranial osteology of Columbidae are not frequent in the literature. This is the first description of the jaw apparatus of G. montana and we had as objectives: to compare its jaw apparatus with other New World Columbidae and thus broaden our anatomical knowledge of the family, and to identify anatomical characters for a future cladistic analysis. We studied 35 specimens from six genera of New World Columbidae. The results showed many structural differences among G. montana and other species, mainly related to the development of some cranial structures, as well as structures particular to some taxa. This possibly indicates that cranial osteology in Columbidae represents a useful tool to systematic studies.
Keywords: Geotrygon montana. Columbidae. Skull. Osteology.
INTRODUÇÃO
Os Columbiformes representam um grupo de aves com ampla distribuição. Provavelmente, originaram-se nas regiões tropicais do Velho Mundo e migraram recentemente para as Américas (há fósseis datando do Terciário, Mioceno na Flórida e Pleistoceno no Brasil) (Sick, 1997). Estão representados no Novo Mundo, apresentando 64 espécies distribuídas em nove gêneros (Howard; Moore, 1998).
Há grande variedade de columbídeos na América do Sul. Ocorrem ao nível do mar, nos Andes, em zonas temperadas, regiões desérticas, bem como em florestas tropicais (Meyer de Schauensee, 1982).
G. montana apresenta dimorfismo sexual no colorido da plumagem, sendo o macho ruivo-purpúreo no dorso e no peito e a fêmea pardo-olivácea; é terrícola, vive no interior da mata ou capoeirões. Ocorre do México a Bolívia, Argentina e Paraguai; e em quase todo o Brasil (Sick, 1997).
Trabalhos sobre a osteologia e miologia cranianas dos Columbiformes são escassos na literatura (Rooth, 1953; Merz, 1963; Andrela; Donatelli, 1995). No que se refere a G. montana, não foram encontrados trabalhos desta natureza, sendo esta, provavelmente, a primeira descrição detalhada da anatomia craniana da espécie.
Muitos autores contribuíram para a sistemática da ordem Columbiformes, entre eles destacam-se Salvadori (1893), Verheyen (1955), Goodwin (1983), Cracraft (1981) e Sibley e Ahlquist (1990), Batista et al. (1997), Johnson et al. (2001) e Livezey e Zusi (2001, 2006 e 2007). Porém, nota-se que, embora seja um grupo reconhecidamente monofilético, o relacionamento da ordem Columbiformes com as demais ordens permanece obscuro e entre seus táxons é controverso nos sistematas.
Sendo assim, procurou-se, neste trabalho, descrever a osteologia craniana em G. montana e, com base nesta descrição, comparar com as demais espécies de Columbiformes do Novo Mundo; estabelecer possíveis padrões anatômicos entre os Columbiformes do Novo Mundo; e ampliar o conhecimento anatômico nesta ordem, bem como identificar caracteres anatômicos para uma futura análise cladística.
MATERIAL E MÉTODOS
Estudou-se a osteologia craniana [O] em 35 espécimens pertencentes a seis gêneros de Columbidae, abrangendo cerca de 70% dos gêneros de Columbiformes do Novo Mundo.
Todo o material faz parte da coleção do Museu Paraense Emílio Goeldi, Belém, Pará, Brasil (MPEG): Geotrygon montana (Linnaeus, 1758) - MPEG: 1758: 261, 619, 665, 1971, 2883, 2884, 2885, 2886, 3309, 3212 [O]; Leptotila rufaxilla (Richard & Bernard, 1792) - MPEG: 1712: 215, 434, 438, 657, 841, 936 [O]; Leptotila verreauxi Bonaparte, 1855 - MPEG: 735 [O]; Patagioenas cayennensis (Bonnaterre, 1792) - MPEG: 92, 399, 920, 921 [O]; Patagioenasspeciosa (Gmelin, 1789) - MPEG: 216, 1972, 3560 [O]; Patagioenas palumbus (Linnaeus, 1758) - MPEG: 214 [O]; Columbina passerina (Linnaeus, 1758) - MPEG: 620, 1031, 3698, 3699 [O]; Columbina squammata (Lesson, 1831); Columbina talpacoti (Temminck,1811) - MPEG: 1139 [O]; Zenaida auriculata (Des Murs, 1847) - MPEG: 3265 [O]; Claravis pretiosa (Ferrari-Perez, 1886) - MPEG: 1635 [O].
As descrições osteológicas detalhadas da espécie Geotrygon montana foram utilizadas como um referencial para as comparações das diversas estruturas anatômicas com as espécies acima mencionadas.
A osteologia craniana foi descrita e desenhada com auxílio de um estereomicroscópio ZEISS Stemi SV11 (ocular 10x e objetivas variando entre 0,6 - 6,6x). Para a mensuração das estruturas ósseas, utilizou-se um paquímetro Mitutoyo com precisão de 0,05 mm.
A nomenclatura para a descrição da osteologia e das estruturas associadas segue a Nomina Anatomica Avium (Baumel et al., 1993).
A nomenclatura e a classificação é aq uela apresentada pelo Comitê Brasileiro de Registros Ornitológicos (CBRO, 2007).
RESULTADOS
OSTEOLOGIA
Ossa cranii
A região do osso frontal (os frontale) (F: Figura 1) limita-se rostralmente com a maxila superior, na região da zona flexoria craniofacialis (ZFCF: Figura 1), onde se articula com duas porções do osso nasal: lateralmente com o processo frontal do nasal (proc. frontalis) (PrFN: Figura 1); e medialmente com o processo frontal do pré-maxilar (proc. premaxilaris) (PrFPM: Figura 1). A região do osso frontal funde-se lateralmente com a região do osso lacrimal sem evidência de sutura e processo lacrimal; medioventralmente funde-se com o septo interorbital e caudalmente funde-se à região parietal. O osso frontal apresenta a superfície dorsal plana, não se observando qualquer depressão frontal. Na região dorsal da órbita, o frontal é mais estreito em relação à região parietal.
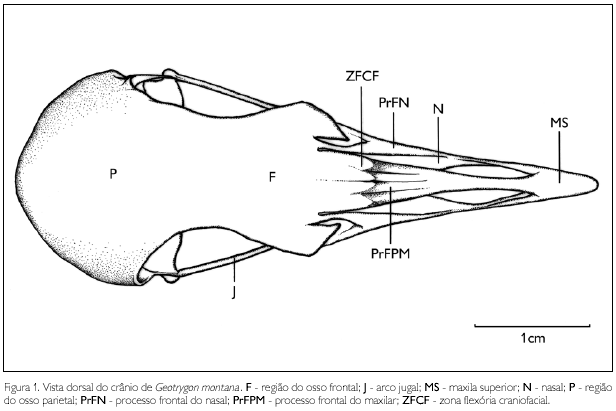
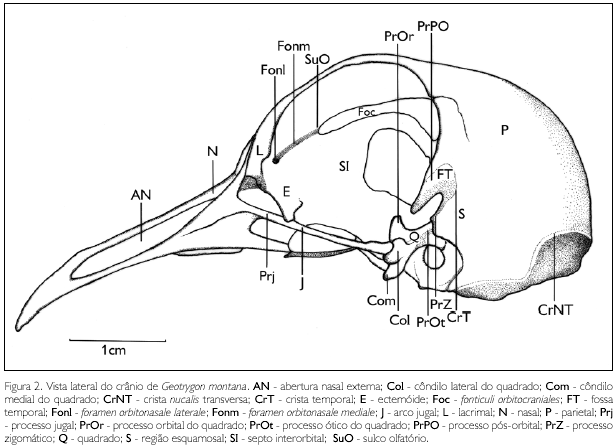
A região parietal (os parietale) (P: Figuras 1 e 2) está confinada à porção caudal do teto craniano. Limita-se caudalmente com a região occipital por meio da crista nucal transversa (crista nuchalis transversa) (CrNT: Figura 2) pouco conspícua e lateralmente com a região esquamosal (S: Figura 2), sendo difícil estabelecer os limites entre estas regiões, haja vista que a crista temporal (CrT: Figura 2), bem como a fossa temporal (FT: Figura 2), em G. montana são pouco conspícuas.
A região occipital compreende as regiões dos ossos supra-occipital, exoccipital e basioccipital fusionados. Observa-se entre estas regiões o forâmen magno (FM: Figura 3), que possui sua abertura desenvolvida em forma circular.
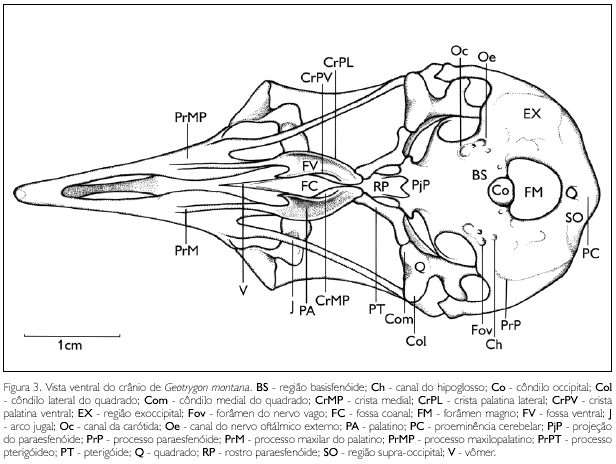
A região supra-occipital (os supraoccipitale) (SO: Figura 3) é delimitada dorsalmente pela crista nucal transversa; funde-se ventrolateralmente com a região exoccipital e ventromedianamente limita-se com o forâmen magno (FM: Figura 3). Observa-se, em sua porção medial, a proeminência cerebelar (proeminentia cerebelaris) (PC: Figura 3) pouco saliente.
A região exoccipital (os exoccipitale) (EX: Figura 3) delimita-se lateralmente com a crista nucal transversa até o processo paraoccipital (proc. paraoccipitalis) pouco desenvolvido; medialmente funde-se com a região basioccipital e laterocaudalmente funde-se à região basisfenóide. Esta espécie não apresenta o tubérculo basilar ou uma crista entre estas regiões, mas destaca-se uma lâmina contínua. Dentre as aberturas encontradas nesta região, destacam-se: canal da carótida (ostium canalis carotici) (Oc: Figura 3); canal do nervo oftálmico externo (ostium canalis ophatalmici externi) (Oe: Figura 3); forâmen do nervo vago (for. nervus vagi) (Fov: Figura 3); e canal do nervo hipoglosso (canalis nervus hipoglossi) (Ch: Figura 3).
Na região basioccipital, o côndilo occipital (condylus occipitalis) (Co: Figura 3) é saliente e a fossa subcond ilar é rasa, pouco conspícua e circunda ventralmente este côndilo.
A região basisfenóide (os basisphenoidale) funde-se rostralmente com a região paraesfenóide e laterocaudalmente às regiões basioccipital e exoccipital.
A lâmina paraesfenóide (os parasphenoidale) é retangular, lisa e afila-se rostralmente na curta projeção paraesfenóide (Pjp: Figura 3), que recobre parte da porção caudal do rostrum parasphenoidale.
O rostro paraesfenóide (rostrum parasphenoidale) (RP: Figura 3) é triangular, estreita-se medialmente formando uma lâmina dorsal que se funde dorsalmente ao septo interorbital e estende-se rostralmente até a porção rostral da fossa coanal do palatino. Na região laterocaudal do rostro paraesfenóide, na face articular pterigóide, destaca-se o curto processo basipterigóide, com ápice reto (que se articula com as projeções encontradas na face medial do osso pterigóide).
O osso ectetmóide (os ectethmoidale) (E: Figura 2) funde-se ao septo interorbital medialmente e ao mesetmóide em sua porção rostromedial e dispõe-se como uma fina lâmina que abrange a porção rostral da órbita. Projeta-se lateralmente e sua extremidade lateral funde-se ao processo orbital do lacrimal, formando o complexo lacrimal-ectetmóide. Sua extremidade laterodorsal também se funde ao osso frontal e apresenta dois foramens: orbitonasalis medial e lateral (for. orbitonasale mediale et laterale) (Fonm e Fonl: Figura 2), sendo o lateral mais desenvolvido que o medial. O osso ectetmóide emite uma projeção lateroventralmente, que atinge a barra jugal, mas sem fundir-se a ela.
O osso mesetmóide (os mesethmoidale) achata-se dorsoventralmente em sua porção dorsal, fundindo-se com a face ventral da região frontal, e sua porção caudal funde-se ao septo interorbital. Ventralmente apresenta-se como uma fina lâmina óssea achatada lateralmente e contínua com o septo interorbital, na porção caudal da abertura anteorbital (fenestra antorbitalis).
O osso lacrimal (os lacrimale) (L: Figura 2) ocupa a porção rostrolateral da órbita. Apresenta o processo supraorbital (proc. supraorbitalis) expandido dorsolateralmente e totalmente fundido à borda lateral da região frontal. Tal processo não se contata com o osso nasal e, portanto, não apresenta uma sutura comum a ambos.
A região esquamosal (os squamosum) (S: Figura 2) limita-se ventrocaudalmente com a cavidade timpânica (meatus acusticus externus); e rostralmente com a região laterosfenóide, por meio do processo pós-orbital (PrPO: Figura 2). A fossa temporal (fossa temporalis) (FT: Figura 2) é pequena, rasa e restrita à porção rostroventral da região esquamosal, ventrocaudalmente ao processo pós-orbital. Esta região possui, ainda, uma discreta concavidade que está confinada à parte lateral do crânio. O processo zigomático é pouco conspícuo (PrZ: Figura 2) e o suprameático não observado.
Orbita
A parede dorsocaudal da órbita recebe contribuições dos ossos orbitoesfenóide (os orbtosphenoidale) e lateroesfenóide (os laterosphenoidale), que se encontram totalmente fundidos nesta região. Limita-se lateralmente com a região esquamosal, medialmente com o forâmen óptico e com o fontículo orbitocranial. Funde-se ventralmente à região paraesfenóide, onde apresenta o conspícuo forâmen maxilomandibular (for. maxillomandibularis). A area muscularis aspera é plana, não se observando elevações e concavidades que possam estar relacionadas à musculatura.
O processo pós-orbital (proc. postorbitalis) (PrPO: Figura 2) localiza-se na porção caudoventral da órbita, é curto e achatado rostrocaudalmente, sendo mais largo em sua base, e afila-se ventralmente.
O septo interorbital funde-se rostrodorsalmente com a face medioventral da região frontal, onde se observa o curto sulco do nervo olfatório (sulcusn. olfactorii) (SuO: Figura 2); caudalmente funde-se à porção mediana da região lateroesfenóide, sendo que, rostralmente a esta fusão, observa-se o forâmen óptico (for. opticum) bem desenvolvido e em forma circular; e ventralmente funde-se ao rostro paraesfenóide. Observa-se que em sua face dorsal, ventralmente à região frontal, o septo interorbital possui uma abertura orbitocranial (fonticuli orbitocraniales) (Foc: Figura 2), que se estende caudalmente até atingir a região orbitoesfenóide, onde se alarga.
Facies
Ossa maxillae et palati
A maxila superior é formada pelos ossos nasal (os nasale), maxilar (os maxillare) e premaxilar (os premaxillare). Funde-se ventrocaudalmente à barra palatina, dorsocaudalmente à região frontal e laterocaudalmente ao arco jugal. Em G. montana, a maxila superior não está disposta em um mesmo plano que o crânio, orienta-se ventralmente, com um ângulo ventral de aproximadamente 40° em relação ao crânio. É longa e estreita, atingindo cerca de 50% do comprimento total do crânio. Seu ápice apresenta curvatura ventral. Os maxilares não se encontram totalmente fundidos entre si, possuem uma abertura ventromedial na maxila contínua com a fenestra palatina (fenestra palatina). Estes ossos fundem-se totalmente aos ossos nasais na porção ventrocaudolateral da maxila superior e no processo jugal (proc. jugalis) (Prj: Figura 2) bem desenvolvido. Os processos maxilopalatinos (proc. maxillopalatinum) (PrMP: Figura 3) são laminares, achatados lateralmente, afilando-se em sua extremidade caudal. Localizam-se na região mediocaudal da maxila superior e estão próximos entre si na linha mediana da maxila superior. Os ossos nasais apresentam dois processos estreitos e longos: o processo maxilar (proc. maxillaris) (PrM: Figura 3), que se funde ao osso maxilar em sua porção ventral; e o processo frontal (proc. frontalis), que se funde com o frontal em sua extremidade caudal e possui duas fendas laterais resultantes da não ossificação desta região. A abertura nasal externa (apertura nasi ossea) (AN: Figura 2) é ampla e estreita, abrangendo a maior parte da face dorsal da maxila. Não há septo nasal (septum nasi ossea) evidente separando as duas aberturas nasais. A porção rostral da maxila (rostrum maxillare), onde se encontram os ossos pré-maxilares (os premaxillare), está fundida com uma leve curvatura ventral na extremidade rostral.
O palatino (os palatinum) (PA: Figura 3) articula-se dorsocaudalmente com o rostro paraesfenóide por meio da lamela do palato (lamella palatina); caudalmente com o pterigóide por meio do curto e largo processo pterigóideo (proc. pterygoideus) (PrPT: Figura 3). A lamela palatina emite projeções dorsais (crista dorsolateralis) na zona flexoria parasphenoidale, que se articulam com a face ventrolateral do rostro paraesfenóide. Rostralmente o palatino funde-se à maxila superior por meio dos processos maxilares do palatino (proc. maxillaris) (PrM: Figura 3), extremamente estreitos e delgados, longos e distantes entre si. Estes se alargam discretamente na porção caudal, formando um discreto ângulo caudolateral (angulus caudolateralis) arredondado; a fossa ventral (fossa ventralis) (FV: Figura 3) é delimitada pela crista ventral (crista ventralis) (CrPV: Figura 3) e lateral (crista lateralis) (CrPL: Figura 3), sendo esta última pouco conspícua. As cristas ventralis são salientes e afastadas entre si com ápice rostral pontiagudo. As cristas mediais (crista medialis) (CrMP: Figura 3), além de delimitarem a fossa coanal (fossa choanalis) (FC: Figura 3) profunda e larga, não se unem caudalmente, evidenciando parte da face ventral da extremidade rostral do rostro paraesfenóide.O pterigóide (os pterygoideum) (PT: Figura 3) é estreito, curto e curvado. Articula-se, em sua face medial, com o processo basipterigóide do rostro paraesfenóide. Articula-se rostroventralmente com o palatino por meio da pouco desenvolvida parspalatina. Em sua porção caudal, o pterigóide expande-se e articula-se com duas regiões do osso quadrado: medialmente com o côndilo medial; e dorsalmente com a base medioventral do processo orbital do quadrado. O pterigóide de G. montana apresenta sua face dorsal lisa, evidenciando um bem desenvolvido processo dorsal do pterigóide.
O vômer (os vomer) (V: Figura 3) é uma lâmina longa achatada lateralmente. Caudalmente contata-se com o rostro paraesfenóide e com a lamela dorsal palatina, bifurcando-se lateralmente. Sua face rostral é afilada, atingindo a linha mediana entre os processos maxilopalatinos; nesta porção o vômer é livre, ou seja, não se contata com os ossos da maxila superior.
O arco jugal (arcus jugalis) (J: Figuras 1, 2 e 3) é constituído pelos ossos quadradojugal (os quadratojugale) e jugal (osjugale) fundidos. O arco jugal é fino, delgado, comprido e inclinado obliquamente em relação ao crânio. Funde-se rostralmente ao processojugalis do osso maxilar; caudalmente articula-se, por meio do côndilo quadradojugal (condylus quadratojugalis), com a cótila quadrática (cotyla quadraticus) do côndilo lateral do processo mandibular do osso quadrado.
O osso quadrado (os quadratum) (Q: Figuras 2 e 3) apresenta uma parte central, o corpus quadrati, que emite três processos: ótico, orbital e mandibular. O ótico (proc. oticus) (PrOt: Figura 2) encontra-se bem desenvolvido, estendendo-se obliquamente em direção ao meato acústico externo e medialmente em relação ao processo suprameático, observando-se neste processo dois capítulos: ótico (capitulum oticum), que se articula com a parede dorsal da cavidade timpânica e é mais desenvolvido que o capítulo esquamosal; e esquamosal (capitulum squamosum), que se articula medialmente em relação à base do processo suprameático; entre estes capítulos destaca-se um pequeno sulco. O orbital (proc. orbitalis) (PrOr: Figura 2) é curto e largo, estende-se rostromedialmente, afilando-se em sua extremidade rostral. O mandibular (proc. mandibularis) apresenta dois côndilos: lateral (condylus lateralis) (Col: Figuras 2 e 3), que se articula com a cótila lateral da mandíbula, possui sua face ventral afilada, em forma de 'cunha'; e côndilo medial (condylus medialis) (Com: Figuras 2 e 3), que se articula com a cótila medial da mandíbula e com o pterigóide, onde apresenta uma pequena depressão para articulação, possui sua face ventral comprimida lateralmente em forma de 'cunha', sendo maior que o côndilo lateral. Entre os côndilos, observa-se o sulcus intercondylaris conspícuo. G. montana não apresenta os côndilos caudal e pterigóide no osso quadrado. Na face medial (facies tympanica) do corpus quadrati, observa-se uma pequena depressão (sulcus pneumaticus) rasa.
Ossa mandibulae
A mandíbula é formada pelos ossos articulare, angulare, suprangulare, dentale, spleniale e prearticulare. Porém, percebe-se que em G. montana o osso dentário não está fundido aos demais ossos, apresentando uma sutura característica. Nota-se que o dentário emite duas projeções caudais; uma dorsal mais curta e larga e uma ventral mais estreita e longa.
A mandíbula pode ser subdividida em partes sinfisial, intermediária e caudal. Quanto às suas dimensões, percebe-se que a mandíbula é comprida e estrei ta, sen do qu e a largura máxima en tre os ramos mandibulares atinge somente cerca de 25% do comprimento total da mandíbula.
A pars symphisialis da mandíbula reúne os dois ramos mandibulares. Esta parte é curta, atingindo somente cerca de 15% do comprimento total da mandíbula. Os ramos mandibulares são estreitos em sua porção rostral e alargam-se em sua porção medial e caudal.
A região do osso dentário abrange cerca de metade do comprimento total dos ramos mandibulares, inicia-se na sínfise mandibular até o ângulo mandibular (angulus mandibulae) (AM: Figura 4), localizado na face dorsal da parte intermediária com curvatura ventral bastante acentuada.
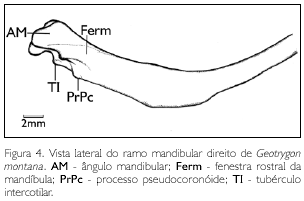
Na parte caudal, os ramos mandibulares, além de largos, são mais espessos. O processo pseudocoronóide (proc. pseudocoronoideus) (PrPc: Figura 4) é pouco conspícuo e localiza-se na porção rostral da cótila lateral. Na face lateral, observa-se uma fina lâmina óssea no local da fenestra mandibular, a qual não está completamente ossificada na porção rostral desta fenestra, evidenciando a fenestra rostral da mandíbula (fenestra rostralis mandibulae) (Ferm: Figura 4) com abertura bastante reduzida. Na face medial há uma fossa (fossa additus canalis mandibulae) extremamente rasa e pouco conspícua, assim como uma fossa lateral (fossa lateralis mandibulae). Já o tubérculo pseudotemporal (tuberculum pseudotemporalis) é pouco conspícuo.
Na face dorsal da porção caudal da mandíbula, constata-se a fossa articular (fossa articularis quadratica) delimitada pela cótila lateral (cotyla lateralis) e pela cótila medial (cotyla medialis) profunda. Estas cótilas estão dispostas de maneira peculiar em G. montana: a cótila lateral dispõe-se rostralmente e é ampla e rasa; já a cótila medial dispõe-se caudalmente à fossa lateral e é mais profunda e estreita. Entre as cótilas, destaca-se o discreto tubérculo intercotilar (tuberculum intercotylaris) (TI: Figura 4) comprimido lateralmente em forma de 'cunha'. Uma característica peculiar de G. montana refere-se à disposição praticamente perpendicular das cótilas em relação aos ramos mandibulares.
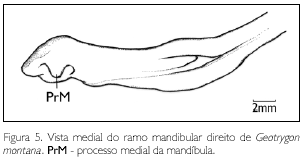
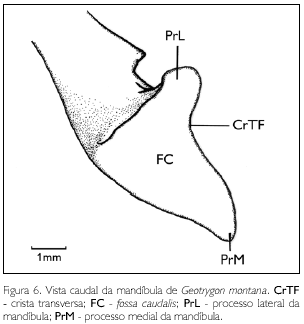
Em sua face medial, a mandíbula apresenta o processo medial (proc. medialis) (PrM: Figuras 5 e 6) curto e pouco desenvolvido, projetando-se medialmente. Em sua face lateral, encontra-se o processo lateral (proc. lateralis) (PrL: Figura 6) bem desenvolvido, com ápice dorsal pontiagudo. Em sua face caudal, observa-se a fossa caudal (fossa caudalis) (FC: Figura 6) rasa e larga (FC) e sua respectiva crista transversa (crista transversa fossae) (CrTF: Figura 6) conspícua. O vértice do angular é ausente em G. montana.
DISCUSSÃO
As características gerais de osteologia craniana que parecem ser comuns a vários Columbidae (Rooth, 1953; Merz, 1963, Gennip, 1986, Andrela; Donatelli, 1995) são: região do osso frontal convexa; articulação com a região da maxila superior, na porção da zona flexoria craniofacialis, por meio de dois processos, frontal do maxilar e nasal do premaxilar; o estreitamento lateral da região frontal em relação à região parietal; a fusão das regiões frontal e lacrimal; a crista e a fossa temporal são pouco conspícuas e quando não o são apresentam uma fossa temporal reduzida; a formação de uma lâmina paraesfenóide na base do crânio; uma curta projeção paraesfenóide; a curta projeção lateroventral do ectetmóide; o curto processo pós-orbital que atinge cerca de 20% da distância total entre a sua origem no crânio e o arco jugal; maxila superior apresenta cerca de 50% do comprimento total do crânio; processos maxilares do palatino longos e estreitos; vômer apresenta-se como uma lâmina longa achatada lateralmente; processo orbital do quadrado curto e alargado; dois côndilos (medial e lateral) no processo mandibular do quadrado, sendo que o côndilo caudal ou posterior funde-se com o côndilo lateral do quadrado (Bock, 1960); e fossa caudal da mandíbula apresenta-se rasa.
Destaca-se em G. montana um processo paraoccipital característico, como observado por Gennip (1986) estudando Patagioenas livia, mas não descrito em C. squammata (Andrela; Donatelli, 1995). A presença de um processo basipterigóide é considerada uma característica de aves mais primitivas, como observou Silveira (1999), e foi descrita em Columbidae apenas em G. montana. O pterigóide de G. montana distingue-se daquele de C. squammata (Andrela; Donatelli, 1995) por ser estreito e curvado; nesta última, o mesmo é curto e largo, sem evidências de curvatura. O processo ótico do quadrado em G. montana e em C. livia (Gennip, 1986) é bem desenvolvido, ao contrário daquele observado em C. squammata (Andrela; Donatelli, 1995). O côndilo medial do quadrado é o mais desenvolvido entre os côndilos do processo mandibular do osso quadrado em G. montana, mas o lateral apresentou-se mais desenvolvido em C. squammata. A pars symphysialis mandibulae apresenta cerca de 1/3 do comprimento total da mandíbula em C. squammata e cerca de 1/4 em G. montana, enquanto o processo medial da mandíbula apresenta-se mais desenvolvido em C. squammata.
O processo zigomático (=esquamosal, Höfling; Gasc, 1984) é pouco conspícuo em G. montana e inexistente em C. squammata, já o processo suprameático é inexistente em ambas; o mesmo foi descrito em C. livia por Gennip (1986). O processo ótico do quadrado em G. montana é similar ao de C. livia descrito e difere daquele de C. squammata; já o processo orbital do quadrado possui morfologia distinta nessas três espécies, sendo mais desenvolvido em C. livia.
Nas demais espécies de Columbidae analisadas, observaram-se as seguintes semelhanças e diferenças com G. montana. Em L. rufaxilla e L. verreauxi, a cótila lateral é rasa e praticamente inconspícua, o côndilo lateral é reduzido e plano, a fossa ventral palatina é mais profunda e os processos maxilopalatinos são mais largos e longos e mais próximos entre si; em Z. auriculata, a cótila lateral é rasa e praticamente insconspícua, o processo retroarticular está presente, sendo mais longo e projetado caudolateralmente, já a fossa ventral palatina é profunda e o côndilo medial do quadrado é menor que em G. montana. Em Claravispretiosa, a cótila lateral é também rasa e insconspícua, como observado nas espécies do gênero Leptotila e em Zenaida auriculata; o processo retroarticular é conspícuo e projetado caudolateralmente; o processo maxilopalatino é tão largo quanto aqueles das espécies de Leptotila, mas próximos entre si. Em Patagioenas speciosa, Patagioenas cayennensis e Patagioenas palumbus, o processo maxilopalatino é o mais amplo dentre todas as espécies estudadas, isto é, mais largo, quase se contatando medialmente, e a fossa ventral palatina é a mais profunda; o processo retroarticular é saliente, porém, menor que em Zenaida auriculata e Claravis pretiosa. Em Columbina passerina, C. squammata e C. talpacoti, os pterigóides são proporcionalmente mais espessos, a fossa ventral palatina mostrou-se tão profunda quanto a de Zenaida auriculata, o processo retroarticular é longo como em Z. auriculata e C. pretiosa; já a projeção caudal do processo maxilopalatino é mais longa que aquela observada nas demais espécies.
Apesar de diversas semelhanças nas estruturas cranianas dos Columbidae terem sido apontadas anteriormente, inúmeras diferenças foram constatadas entre os gêneros analisados, principalmente relativos ao grau de desenvolvimento; todavia, há estruturas que se apresentam em alguns gêneros, como o processo retroarticular e a projeção caudal palatina, as quais distinguem os gêneros de maneira mais efetiva que o simples desenvolvimento. Um estudo mais profundo das espécies de Columbidae talvez venha a revelar estruturas características de um determinado gênero de tal forma que se possa caracterizá-lo. Isso indicaria que a osteologia craniana dos Columbidae pode ser uma ferramenta importante para estudos sistemáticos.
CONCLUSÕES
De uma forma geral, G. montana difere-se em alguns aspectos da osteologia craniana se comparada com outras espécies de Columbidae descritas por outros autores:
1) Leptotila rufaxilla, L. verreauxi: a cótila lateral é rasa e praticamente inconspícua, o côndilo lateral é reduzido e plano, a fossa ventral é mais profunda e os processos maxilopalatinos são mais largos, próximos entre si e os mais longos encontrados.
2) Zenaida auriculata: a cótila lateral é rasa e praticamente insconspícua, o processo retroarticular é mais longo e projetado caudolateralmente, a fossa ventral é mais profunda, a fossa temporal é mais reduzida e rasa, o côndilo medial do quadrado é menor que em G. montana.
3) Claravis pretiosa: a cótila lateral é rasa e praticamente insconspícua, o processo retroarticular é mais longo e projetado caudolateralmente, os processos maxilopalatinos são mais largos e próximos entre si, a fossa temporal é bastante reduzida e o processo pós-orbital é espicular e reduzido.
4) Patagioenas speciosa, Patagioenas cayennensis, Patagioenas palumbus: o processo maxilopalatino é amplo (mais largo, quase se contatando medialmente), fossa ventral mais profunda, o tubérculo basilar é mais elevado, a cótila lateral é mais larga e rasa, o processo retroarticular é mais saliente, porém, menor que Zenaida e Claravis.
5) Columbina passerina, C. squammata, C. talpacoti: pterigóides proporcionalmente mais espessos, fossa ventral mais profunda, lacrimal mais estreito dorsolateralmente (pouco ultrapassa a linha lateral do arco jugal), o processo retroarticular é longo como em Zenaida e Claravis, a projeção caudal do processo maxilopalatino é mais longa.
AGRADECIMENTOS
Ao Sr. Elielson Rocha, pela confecção das ilustrações e aos dois revisores anônimos pelas críticas e sugestões.
REFERÊNCIAS
ANDRELA, S.; DONATELLI, R. J. Osteología e miologia cranianas de Columbina squammata (Lesson, 1831) (Aves:Columbiformes). Naturalia, São Paulo, v. 20, p. 107-23, 1995.
BATISTA, L. F.; TRAIL, P. W.; HORBLIT, H. M. Family Columbidae (Pigeons and Doves). In: HOYO, J. del; ELLIOT A.; SARGATAL, J. (Eds.). Handbook of the birds of the world. Barcelona: Lynx Ed., 1997. v. 4. p. 60-245. Sandgrouse to Cuckoos.
BAUMEL, J.; KING, A. S.; BREAZILE, J. E.; EVANS, H. E.; VANDEN BERGE, J. C. Handbook of avian anatomy: nomina anatomica avium. 2. ed. Cambrige: Nuttall Ornithological Club, 1993.
BOCK, W. J. Secondary articulation of avian mandibule. Auk, Washington, v. 77, n. 1, p. 19-55, 1960.
CRACRAFT. Toward a phylogenetic classification of the recent birds of the word (Class Aves). Auk, Washington, v. 98, n. 4, p. 681-714, 1981.
CBRO. Comitê Brasileiro de Registros Ornitológicos. Disponível em: <http://www.cbro.org.br>. Acesso em: 31 abr. 2007.
GENNIP, E. M. S. J. van. The osteology, arthrology and myology of the jaw apparatus on the pigeon (Patagioenaslivia L.). Neth. J. Zool., Wageningen, v. 36, n. 1, p. 1-46, 1986.
GOODWIN, D. Pigeons and doves of the world. 3rd ed. New York: Cornell University Press, 1983. 446 p.
HÖFLING, E.; GASC, J. P. Biomécanique du crâne et du bec chez Ramphastos (Aves, Ramphastidae). I. Description des éléments anatomiques. Gegenbaurs Morphol. Jahrb., Leipzig, v. 130, n. 1, p. 125-47, 1984.
HOWARD, R.; MOORE, A. A complete checklist of the birds of the world. 2nd ed. San Diego: Academic Press, 1998. 630 p.
JOHNSON, Kevin P.; KORT, Selvino de; DINWOODEY, Karen; MATEMAN, A. C.; TEN CATE, Carel; LESSELLS, C. M.; CLAYTON, Dale H. A molecular phylogeny of the dove genera Streptopelia and Columba. Auk, Washington, v. 118, n. 4, p. 874-87, 2001.
LIVEZEY, B. C.; ZUSI, R. L. Higher-order phylogenetics of modern Aves based on comparative anatomy. Neth. J. Zool., Wageningen, v. 51, p. 179-206, 2001.
LIVEZEY, B. C.; ZUSI, R. L. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy: I. - Methods and characters. Bull. Carnegie Mus. Nat. Hist., v. 37, p. 1-544, 2006.
LIVEZEY, B. C.; ZUSI, R. L. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy: II. - Analysis and discussion. Zool. J. Lin. Soc., London, v. 149, p. 1-94, 2007.
MERZ, R. L. Jaw musculature of the mourning and White-winged Doves. Univ. Kans. Publ. mus. Nat. Hist., Lawrence, v. 12, p. 521-51, 1963.
ROOTH, J. On the correlation between the jaw muscles and the structure of the skull in Patagioenaspalumbuspalumbus. L. Proc. K. Ned Akad. Wet., Amsterdan, v. 56, p. 251-64, 1953.
SALVADORI, T. Catalogue of birds in British Museum. London: Britsh Museum (Natural History), 1893. v. 21.
MEYER de SCHAUENSEE, R. A guide to the birds of South America. Philadelphia: Academy of Natural Sciences of Philadelphia, 1982. Reimpressão de 1970.
SIBLEY, C. G.; AHLQUIST, J. E. Phylogeny and classification of birds. New Haven: Yale Univ. Press, 1990. 1111 p.
SICK, H. Ornitologia brasileira. Rio de Janeiro: Nova Fronteira, 1997. 862 p.
SILVEIRA, L. F. Osteologia craniana dos Tinamidae (Aves: Tinamiformes), com implicações sistemáticas, 1999. 109 p. Dissertação (Mestrado) - Instituto de Biociências, Universidade de São Paulo, São Paulo, 1999.
VERHEYEN, R. Contribution à la systématique des Piciformes basée sur l'anatomie comparée. Bull. Inst. R. Sci. Natur, Belg., Bruxelles, v. 31, p. 1-43, 1955.
 Endereço para correspondência:
Endereço para correspondência:
Museu Paraense Emílio Goeldi
Editor do Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais
Av. Magalhães Barata, 376
São Braz - CEP 66040-170
Belém - PA - Brasil
Caixa Postal 399
Telefone/fax: 55-91- 3249 -1141
E-mail: boletim@museu-goeldi.br
Recebido: 05/03/2004
Aprovado: 13/07/2007
