










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.2 n.3 Belém dez. 2007
As florestas de Belo Monte na grande curva do rio Xingu, Amazônia Oriental
The forests of Belo Monte on the great curve of the Xingu River, Eastern Amazon
Rafael de Paiva SalomãoI; Ima Célia Guimarães VieiraII; Chieno SuemitsuIII; Nélson de Araújo RosaIV; Samuel Soares de AlmeidaV; Dário Dantas do AmaralVI; Moirah Paula Machado de MenezesVII
IMuseu Paraense Emílio Goeldi. Belém, Pará, Brasil (salomao@museu-goeldi.br)
IIMuseu Paraense Emílio Goeldi. Belém, Pará, Brasil (ima@museu-goeldi.br)
IIIUniversidade Federal do Pará. Campus Santarém. Centro de Ciências Biológicas. Belém, Pará, Brasil (chieno@ufpa.br)
IVMuseu Paraense Emílio Goeldi. Belém, Pará, Brasil
VMuseu Paraense Emílio Goeldi. Belém, Pará, Brasil (samuel@museu-goeldi.br)
VIMuseu Paraense Emílio Goeldi. Belém, Pará, Brasil (dario@museu-goeldi.br)
VIIUniversidade Federal do Pará. Belém, Pará, Brasil (moirah@ufpa.br)
RESUMO
No intuito de se caracterizar a flora da região de Belo Monte ou Volta Grande do Xingu, efetuou-se uma análise da florística, fitossociologia, estrutura e etnobotânica nas fitofisionomias florestais na área de estudo que inclui os territórios de Altamira, Anapu, Senador José Porfírio e Vitória do Xingu, nas mesorregiões do baixo e médio Xingu, no estado do Pará. Nas quatro fitofisionomias florestais majoritárias (floresta ombrófila densa, floresta ombrófila aluvial, floresta ombrófila aberta com palmeira, floresta ombrófila aberta com cipó e palmeira), foram lançadas parcelas de estudos que abrangeram uma área total de 24,3 ha. Foram amostrados indivíduos em três categorias de diâmetros minímos de inclusão: DAP ≥ a 5, 10 e 30 cm. Registrou-se um total de 13.790 indivíduos compreendendo 662 espécies, distribuídas em 65 famílias botânicas. Comparando-se as fitofisionomias estudadas, concluiu-se que a floresta ombrófila densa tem a maior riqueza de espécies (433), seguindo-se a floresta ombrófila aberta com cipó e palmeira (264), a floresta ombrófila aluvial (203) e a floresta ombrófila aberta com palmeira (140). Com relação às espécies raras, a floresta ombrófila densa apresentou o maior número (141), em contraposição à floresta ombrófila aberta com palmeira, que apresentou o menor (63). Caesalpiniaceae foi a família que apresentou o maior índice de valor de importância (1VIF) e de cobertura (IVCF) em todas as fitofisionomias estudadas, à exceção da floresta ombrófila aberta com cipó e palmeira (Lecythidaceae). Alexa grandiflora e Voucapoua americana foram as espécies de maiores índices (1V1 e 1VC) em todas as fitofisionomias estudadas, exceto na floresta ombrófila aluvial (Pterocarpus amazonicus e Molia luscens). Estimou-se para todos os ecossistemas analisados um total de 92,68 km2 de cobertura florestal em toda a área de estudo. Estimou-se para esta área uma abundância total de 403.069.870 árvores com DAP ≥ 10 cm, volume total de madeira de 196.276.924 m3 e uma biomassa aérea viva de 198.503.191 t. Foram identificadas algumas espécies florestais de grande interesse para os programas de conservação da flora, como uma espécie arbórea praticamente extinta nas áreas onde é possível a penetração humana, o 'pau cravo' (Dicypellium caryophyllatum), uma Lauraceae muito cobiçada pela indústria perfumista; e uma outra espécie de difícil ocorrência (Sagotia brachysepala), uma Euphorbiaceae de dispersão muito restrita.
Palavras-chave: Floresta pluvial tropical. Florística. Estrutura. Belo Monte. Fitofisionomia. Etnobotânica.
ABSTRACT
In order to characterize the forests of the Belo Monte region at the 'Volta Grande' of the Xingu River, Brazil, a floristic, phytosociological, structural and ethno-botanical analysis was undertaken in forested areas of Altamira, Anapu, Senador José Porfirio, and Vitoria do Xingu, in the Lower and Middle Xingu River regions of Fará. In the four most common forest formations (dense broadleaf forest, broadleaf flood forest, open broadleaf forest with palms, and broadleaf forest with lianas and palms) study plots were laid out that covered a total area of 24.3 ha. Surveys included individuals in three diameter size classes: DBH ≥ 5, 10, and 30 cm. A total of 13.790 individual plants was recorded, comprising 662 species distributed in 65 botanical families. In comparing the forest types, it is concluded that the dense broadleaf forest is the most species rich (433 spp.), followed by the open broadleaf forest with lianas and palms (264 spp.), the broadleaf flood forest (203 spp.), and the open broadleaf forest with palms (140 spp.). In relation to rare species, the dense broadleaf forest showed the greatest number (141 spp.), in contrast to the open broadleaf forest with palms that showed the least (63 spp.). Caesalpiniaceae was the family that had the greatest indices of economic importance (IVIF) and oftotal coverage (IVCF) in all of the studied forest types except for the open broadleaf forest with lianas and palms (Lecythidaceae). Alexa grandiflora and Voucapoua americanawere the species with the greatest indices (economic importance and total coverage) in all the studied Forest formations, except in the broadleaf flood forest (Pterocarpus amazonicus and Molia luscens) . It is estimated that these analyzed ecosystems have a combined area of 92,68 km2 in the study area. A total abundance of 403,069,870 trees with DBH ≥ 10 cm is estimated, with a total wood volume of 196,276,924 m3 and a live aerial biomass of 198,503,191 metric tons. Forest species of great interest for floral conservation programs were identified, such as the practically extinct Amazon cinnamon tree 'pau cravo' (Dicypellium caryophyllatum) , a member of the Lauraceae that is much sought-after by the perfume industry, and another rarely found species (Sagotia brachysepala) , a member of the Euphorbiaceae with a very restricted geographical distribution.
Keywords: Tropical rain forest. Forest composition. Forest structure. Belo Monte. Phytophysionomy. Ethnobotany.
INTRODUÇÃO
A bacia amazônica é a maior bacia hidrográfica do mundo, com uma drenagem de cerca de 6 milhões de km2, sendo 3,9 milhões no Brasil. Entre os principais afluentes da margem esquerda encontram-se os rios Japurá, o Negro e o Trombetas; na margem direita, o Juruá, o Purus, o Madeira, o Xingu e o Tapajós. Esses tributários, devido às características de seu perfil topográfico, contendo setores de alternância brusca de altitudes, apresentam i nteresses em termo de aproveitamento hidrelétrico, como o caso do rio Xingu (Projeto Radam, 1974).
O rio Xingu pertence ao grupo dos rios de águas claras, pobres em sedimentos, mas em outras áreas podem ser ricas em matéria dissolvida (Ayres, 1995). Possui uma extensão de 1.500 km, desde suas nascentes n o Planalto Cen tral Brasi leiro até sua foz n o rio Amazonas, e drena uma área de 540. 000 km2, com débito de 7.100 m3/seg (Miranda et al., 1988). Nasce a oeste da Serra do Roncador e ao norte da Serra Azul, no leste do Mato Grosso. Corre na direção sul-norte, paralelo aos rios Tapajós e Tocantins, e após percorrer pouco mais de 2.000 km, deságua ao sul da ilha de Gurupá (Pará), na margem direita do Amazonas, do qual é um dos maiores afluentes.
O Projeto Radam (1974) chamou a atenção em suas recomendações para um fato digno de se salientar em referência a 'Volta Grande do Xingu', onde o nível do rio Xingu desce cerca de 70 m em todo o percurso de Volta Grande, oferecendo um dos maiores potenciais hidráulicos do baixo Amazonas. Tal dádiva natural estaria intrinsicamente relacionada às ocorrências de platôs da Formação Barreiras (potencialidade bauxífera) em Monte Dourado e na região Moju-Capim-Paragominas, bem como a vastíssima extensão dessa formação a oeste do rio Xingu, acrescentando que qualquer implantação de indústrias de alumínio na região demandaria energia elétrica barata, cuja fonte seria, sem dúvida, de origem hidráulica.
A área denominada de Belo Monte ou Volta Grande do Xingu inclui territórios de cinco municípios (Altamira, Anapu, Brasil Novo, Senador José Porfírio e Vitória do Xingu) das mesorregiões do baixo e médio Xingu, noestado do Pará (MPEG, 2002).
O nível das informações científicas sobre os mais diversos aspectos dos ecossistemas amazônicos, como solos, geomorfologia, hidrologia, limnologia, geologia, biogeografia, paleoecologia e ecologia, aumentou consideravelmente nos últimos anos (Pires et al., 1953; Hueck, 1966; Falesi, 1972; Ab'Saber, 1982; Absy, 1982; Haffer, 1982; Sioli, 1984; Whitmore; Prance, 1987; Campbell; Hammond, 1989; Daly; Prance, 1989; Pires; Prance, 1985, Prance; Lovejoy, 1985), embora se reconheça que muito conhecimento científico adicional ainda precisa ser gerado.
A região passou por considerável mudança na sua cobertura florestal original, com a ação antrópica mais fortemente presente na zona de influência da rodovia Transamazônica (BR-230) e suas transversais, cuja ocupação humana tem sido intensamente induzida por projetos de colonização agrária nos últimos 30 anos. Em média, cerca de 60% da cobertura vegetal original foram convertidos em diversos tipos de usos da terra, entre os quais agricultura familiar de ciclo curto, pastagens para pecuária bovina e culturas perenes, como cacau e pimenta do reino. As principais unidades de paisagens são: florestas ombrófilas ou úmidas, podendo ser abertas ou densas; vegetação aluvial, diretamente influenciada pelo sistema hidrológico do rio Xingu; e florestas secundárias recentes e antigas, áreas de capoeiras abandonadas pela agricultura familiar e uso agropecuário, que agrupa áreas de culturas de ciclos curto, longo e pastagens (MPEG, 2002).
No intuito de se conhecer a flora das diversas fitofisionomias florestais primárias da região de Belo Monte, na grande curva do Xingu, efetuou-se uma análise da vegetação florestal em relação à florística, fitossociologia, estrutura e etnobotânica das diversas fitofisionomias florestais presentes na área de estudo.
MATERIAL E MÉTODOS
ÁREA DE ESTUDO
A área denominada de Belo Monte inclui territórios de 11 municípios (Altamira, Anapu, Brasil Novo, Gurupá, Medicilândia, Pacajá, Placas, Porto de Moz, Senador José Porfírio, Uruará e Vitória do Xingu) das mesorregiões do baixo e médio Xingu, todos no estado do Pará. Os levantamentos biológicos da vegetação florestal deste estudo abrangeram os quatro municípios mais importantes desta região, também conhecida como a 'grande curva do Xingu': Altamira, Anapu, Vitória do Xingu e Senador José Porfírio. Cerca de 250 mil pessoas vivem na região que tem como elemento integrador a Transamazônica e o Xingu, em sua parte navegável, e Altamira (maior município brasileiro em área física) como o mais relevante centro urbano regional, com cerca de 65.000 habitantes (Figura 1).
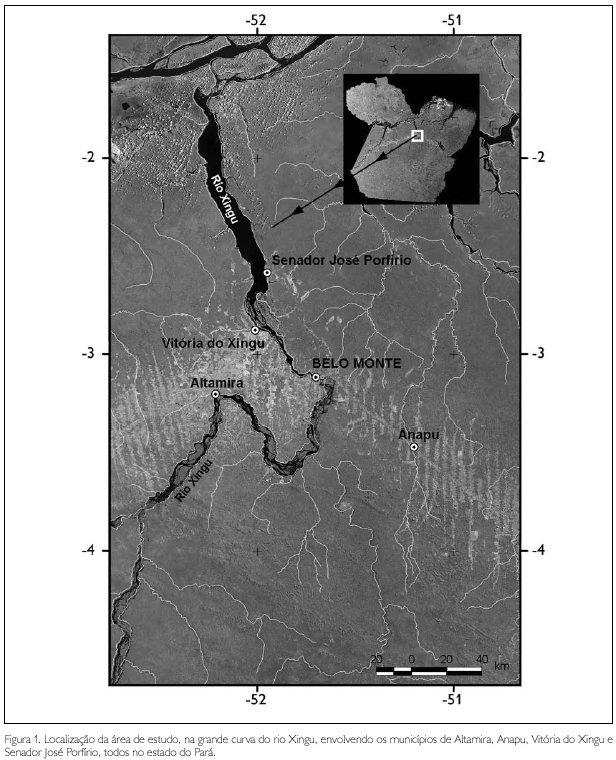
O município de Vitória do Xingu é o que possui área mais extensa de cobertura florestal na região de Belo Monte, seguindo-se Altamira, Senador José Porfírio e Anapu (Tabela 1).
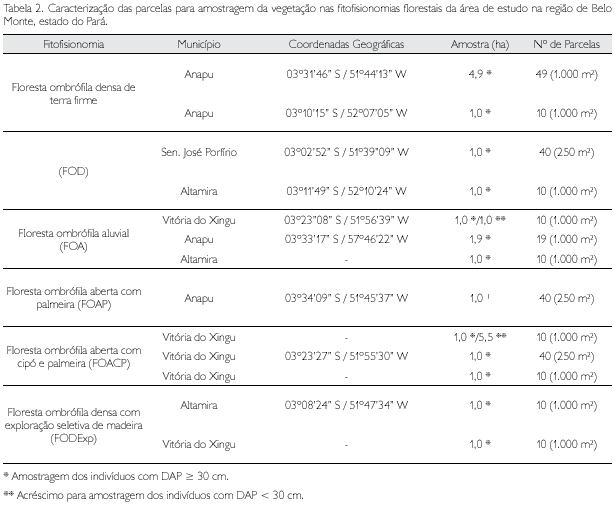
Na área de estudo, foram identificadas quatro tipologias florestais primárias (MPEG, 2002): (a) floresta ombrófila densa de terra firme, com emergentes e relevo acidentado (primária e com extração seletiva de madeira); (b) floresta ombrófila aberta com palmeira e relevo acidentado; (c) floresta ombrófila aberta com cipó e palmeira e relevo acidentado; e (d) floresta ombrófila aluvial periodicamente inundada. Nestas tipologias, que compõem a fitofisionomia majoritária de toda a região estudada, foram implantadas parcelas para amostragem da vegetação florestal (Tabela 2).
AMOSTRAGEM DA VEGETAÇÃO
A escolha dos locais de amostragem baseou-se nas fitofisionomias florestais predominantes na área, cuja identificação primária foi executada por meio da interpretação de imagens orbitais Landsat TM, confirmada através de sobrevôo de reconhecimento na área.
Efetuou-se um reconhecimento terrestre após o sobrevôo pela área para o estabelecimento das parcelas de amostragens. Paralelamente ao levantamento de dados florísticos e estruturais, procedeu-se as coletas botânicas para a elaboração da listagem florística. Foram realizadas duas campanhas de campo.
O material coletado teve o processamento usual de herborização, identificadas preliminarmente e encaminhadas para o Herbário do Museu Paraense Emílio Goeldi (MG) para identificação e incorporação do material fértil ao acervo institucional.
Forma, tamanho e número de parcelas
A forma das parcelas de estudos foi a retangular com dimensões de 20 m x 50 m (DAP ≥ 5 cm e DAP ≥ 30 cm) e de 10 m x 25 m (DAP ≥ 10 cm); o número de parcelas foi variável (Tabela 1).
Para as estimativas dos parâmetros fitossociológicos, as parcelas múltiplas de quatro foram agrupadas em uma única para uniformização da área com as demais parcelas.
Qualificação dos indivíduos amostrados
A amostragem, independentemente da tipologia florestal, obedeceu a três limites mínimos de DAP (diâmetro do tronco a 1,3 m do solo) para a qualificação dos indivíduos amostrados: DAP ≥ 5 cm, DAP ≥ 30 cm e DAP ≥ 10 cm.
No limite mínimo de 5 cm, foram mensurados o DAP e a altura total de cada indivíduo. A altura do fuste foi estimada somente para indivíduos com DAP mínimo de 10 cm, possibilitando, desta forma, a geração de estimativas de volumes de madeira nas diversas fitofisionomias.
Intensidade da amostragem por classe diamétrica
• Diâmetro igual ou superior a 5 e inferior a 10 cm (5 cm < DAP < 10 cm): correspondente ao sub-bosque da floresta → 14,8 ha amostrados;
• Diâmetro igual ou superior a 10 cm (DAP ≥ 10 cm): correspondente à floresta propriamente dita → 17,8 ha;
• Diâmetro igual ou superior a 30 cm (DAP ≥ 30 cm): para estimativas da volumetria de madeira e de biomassa → 24,3 ha (14,8 ha para DAP ≥ 5 cm e 9,5 ha para DAP ≥ 30 cm).
Procedimentos de campo
A medição do diâmetro do fuste (obtida através de fita diamétrica) foi feita sempre a 1,3 m do solo (DAP) ou acima das sapopemas ou bifurcações, irregularidades etc.; usou-se uma vara de alumínio com 1,3 m de comprimento para determinação do local exato de medição. As árvores que se situavam no limite esquerdo da unidade amostral ou parcela (UA), parcialmente dentro da UA e parcialmente fora, foram registradas, enquanto as do limite direito não.
Para as estimativas das alturas do fuste e da copa, utilizou-se uma vara de 5 m de comprimento, marcada de metro em metro. Estas estimativas eram fornecidas sempre pelo mesmo auxiliar de campo.
PARÂMETROS ESTIMADOS
As análises florística, fitossociológica e estrutural da vegetação obedeceram a dois pressupostos metodológicos: a fitofisionomia e os limites mínimos de qualificação do DAP dos indivíduos amostrados. Assim, para cada fitofisionomia definida pelo mapeamento da vegetação, e onde houve amostragem de campo, analisou-se a vegetação para cada um daqueles limites mínimos de DAP.
A florística do ecossistema florestal primário foi caracterizada utilizando-se as famílias e espécies das unidades amostrais. A análise do grau de semelhança entre as UA's foi calculada através do índice de similaridade Sõrensen (1943) e a diversidade florística analisada através do índice de Shannon & Wiener (Shannon; Wiener, 1949; Margalef, 1968; Ricklefs, 1979) e de Simpson (1949) (Tabela 3).
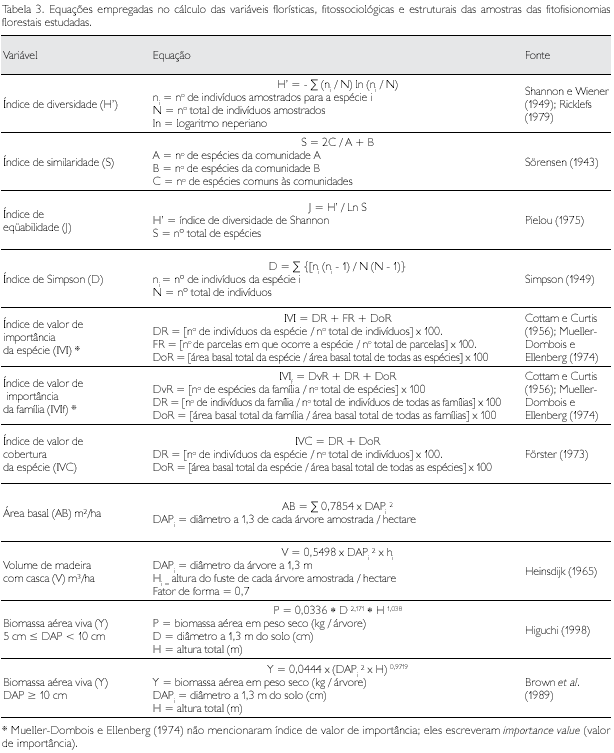
As variáveis fitossociológicas como densidade, diversidade, freqüência e dominância foram calculadas em nível de família e de espécie, possibilitando a avaliação dos índices de importância das famílias e espécies Cottam e Curtis (1956) e Mueller-Dombois e Ellenberg (1974). O número de indivíduos por hectare, a área basal, o volume de madeira e a biomassa aérea viva (Brown et al., 1989, Uhl et al., 1988) também foram estimados (Tabela 3).
PROCEDIMENTOS METODOLÓGICOS
Analisou-se a estrutura da vegetação inerente a cada fitofisionomia com relação à distribuição diamétrica e altimétrica, volumetria de madeira, área basal e produção de biomassa.
Descreveu-se a utilidade mais comum de todas as espécies amostradas, ressaltando-se o valor comercial da madeira, assim como o seu uso alternativo, ou seja, valor dos frutos para o homem e/ou fauna, produção de resina, látex, essência aromática, corante, uso medicinal etc.
Considerou-se floresta propriamente dita, neste estudo, aquela formação vegetal cujos espécimes amostrados têm DAP ≥ 10 cm, e como sub-bosque, o estrato cujos indivíduos estão abaixo deste limite e acima de 5 cm de DAP.
Considerou-se espécie rara aquela que ocorreu com apenas um indivíduo na amostragem relativa a cada fitofisionomia; analogamente, a família que apresentou uma espécie foi considerada rara e, caso esta espécie tenha ocorrido com apenas um indivíduo, foi considerada raríssima.
FITOFISIONOMIAS FLORESTAIS ANALISADAS
Na região de Belo Monte, foram analisadas, neste estudo, a vegetação referente a quatro fitofisionomias da floresta ombrófila: floresta ombrófila densa, floresta ombrófila aluvial, floresta ombrófila aberta com palmeira e floresta ombrófila aberta com cipós e palmeiras; sendo também analisada a floresta ombrófila densa com exploração seletiva de madeira.
As florestas ombrófilas (ou floresta pluvial tropical), conhecidas regionalmente como de terra firme, constituem a formação mais representativa da região amazônica, totalizando cerca de 3,5 milhões de km2 (Pires, 1973). Elas incluem uma grande variedade de subtipologias e ambientes. Essas florestas podem apresentar diferentes fisionomias e paisagens, desde aquelas densas de terras baixas, com elevada biomassa e diversidade, até florestas abertas submontanhosas e montanhosas, de menor porte e riqueza de espécies.
A heterogeneidade específica é uma característica importante dessas florestas, pois mesmo no domínio de uma mesma paisagem, a mudança na composição das espécies varia consideravelmente de local para local (Pires, 1973; Prance, 1979; Pires; Prance, 1985). Outro traço característico dessas florestas é o elevado número de espécies com poucos indivíduos (localmente raras, com 1 indivíduo/hectare) e o reduzido número de espécies dominantes, abundantes e freqüentes (Pires et al., 1953; Almeida et al., 1993).
As florestas ombrófilas, ou úmidas, crescem sobre os mais diferentes tipos de solos e relevos da Amazônia. Os solos e suas propriedades químicas provavelmente não se constituem fatores limitantes às florestas ombrófilas, uma vez que possuem um complexo e fechado sistema de ciclagem de nutrientes que lhes permite viver de si mesmas. Ou seja, toda a demanda nutricional é atendida pela própria fitomassa. Para a ciclagem de nutrientes, essas florestas contam com mecanismos eficientes de fragmentação do material vegetal, decomposição da matéria orgânica e liberação e reabsorção de nutrientes. Nestes sistemas estão presentes associações mutualísticas com insetos e microrganismos.
Na região de Belo Monte estão presentes florestas ombrófilas abertas e densas sobre altos platôs do interflúvio Xingu-Tapajós, ao norte; e as superfícies arrasadas, residuais da serra dos Carajás (RADAMBRASIL, 1976) a oeste e sul. Pontualmente, o domínio das florestas densas compreende a parte oeste à margem direita do rio Xingu, limitada ao sul pelo rio Bacajá. Inclui terrenos de relevo dissecado, residual de antigos platôs, com vales profundos e bem recortados.
Essas florestas são as mais pressionadas pela atividade madeireira, por apresentarem o maior número de espécies de madeira de lei e nobre, a exemplo do mogno (Swietenia macrophylla, Meliaceae), jatobá (Hymenaea courbarl, Caesalpiniaceae), cedro (Cedrela odorata, Meliaceae), maçaranduba (Manilkara huberi, Sapotaceae), ipês (Tabebuia serratifolia, T. impetiginosa, Bignoniaceae), tauari (Couratari multiflora, C. guianensis, Lecythidaceae), dentre outras espécies valiosas (Veloso et al., 1991).
Floresta ombrófila densa - Floresta de terra firme
Este é o tipo de vegetação predominante na Amazônia. As florestas densas abrangem cerca de dois milhões de km2. Estão em toda a bacia Amazônica, geralmente associadas às planícies de terras baixas (até 1 m de altitude), podendo ocorrer também em relevo submontanhoso. Esse tipo de vegetação é o mais representativo na região de Belo Monte, seguido das florestas abertas com palmeiras. Está situado na margem direita do rio Xingu, acima e a oeste da vila de Belo Monte, município de Vitória do Xingu.
Em termos paisagísticos, a exuberância das florestas densas amazônicas é produto da elevada disponibilidade de energia solar, água e solos profundos com boa capacidade de retenção hídrica. Essas florestas são sempre-verdes, latifoliadas e sem estacionalidade marcante. A maior parte dessas florestas cresce sobre latossolo amarelo, amarelo-vermelho eplintossolos, com textura variando de areno-argilosa a argilosa. Os solos, em sua maioria, são profundos e com boa retenção hídrica. O dossel é denso, fechado e compacto, situado entre 30 e 35 m de altura. Nestas condições, o dossel intercepta grande parte da energia solar, passando somente uma pequena fração, em torno de 5%, que chega até o chão da floresta.As características de história natural mais marcantes destas espécies são a dispersão predominante por mamíferos, sementes ou propágulos de tamanho e massa consideráveis, com pouca capacidade de dormência e pequeno tempo de viabilidade. Ao contrário das espécies de capoeira, que formam bancos de sementes, as espécies dessas florestas formam bancos de mudas. As estratégias para a regeneração natural bem sucedida são muitas, inclusive espécies especialistas de clareiras naturais, tolerantes ao sombreamento em diversos graus.
As espécies mais comuns neste estrato são a castanheira (Bertholletia excelsa, Lecythidaceae), melancieira (Alexa grandiflora, Fabaceae), acapu (Vouacapoua americana, Caesalpiniaceae), babaçu (Attalea speciosa, Arecaceae), capoteiro (Sterculia speciosa, Sterculiaceae), seringueira (Hevea brasiliensis, Euphorbiaceae), tachi preto (Tachigalia myrmecophila, Caesalpi n i aceae), amarelão (Apuleia mollaris, Caesalpiniaceae), ingá pereba (Inga alba, Mimosaceae), dentre outras.
Os sub-bosques destas florestas são limpos e sombreados, com dominância de espécies pertencentes às famílias Heliconiaceae, Marantaceae e Strelitziaceae. Nos locais mais úmidos deste estrato, formam-se tapetes de ervas, samambaias e lianas herbáceas. É comum encontrar também palmeiras pequenas, como a mumbaca (Astrocaryum gynacanthum), bacabi (Oenocarpus minor), ubim-açu (Geonoma maxima) e jacitara (Desmoncus spp.).
O estrato emergente é composto por espécies arbóreas que alcançam c.a. 45 m de altura. No geral, são árvores raras, com densidades abaixo de 1 árvore/ hectare. Entre as espécies emergentes mais conspícuas pode-se citar, além da castanheira-do-pará (Bertholletia excelsa, Lecythidaceae), angelim pedra (Hymenologium petraeum, Fabaceae), cu m aru (Dipteryx odorata, Fabaceae), faveira (Parkia oppositifolia, Mimosaceae), melancieira (Alexagrandiflora, Fabaceae) e tauari (Couratari minutiflora, Lecythidaceae).
Na área estudada, este tipo de vegetação está assentada sobre relevos arrasado, residual da Serra dos Carajás e planalto dissecado com vales em forma de 'V'. A topografia subjacente a este tipo vegetacional varia de relevo plano a medianamente ondulado. A malha de pequenos igarapés de terra firme produz uma topografia composta por platôs, baixios e vertentes. As vertentes mais acentuadas têm inclinação em torno de 40o.
Os cipós nas florestas densas não dominam com o em algu m as florestas abertas; em bora se deva registrar a presença de espécies dos gêneros Derris (Fabaceae), Machaerium (Fabaceae), Moutabea (Polygalaceae), Bauhinia (Caesalpiniaceae) e das famílias Bignoniaceae e Dilleniaceae.
As palmeiras aparecem nas áreas mais altas, com o babaçu (Attalea speciosa), e nos baixios, onde o solo é mais úmido e arenoso. Nestes locais, elas são representadas pela bacaba (Oenocarpus bacaba), paxi ú ba (Socratea exorrhiza) e, esporadicamente, o patauá (Oenocarpus bataua).
A diversidade das florestas amazônicas densas é uma das mais elevadas da região neotropical, variando de 115 a 240 espécies de árvores com DAP ≥ 10 cm. A distribuição de abundância de indivíduos nestas florestas registra um elevado número de espécies localmente raras (c.a. 50%), com 1 árvore/hectare e outro grupo restrito de espécies, com elevado número de indivíduos.
Floresta ombrófila aluvial - Florestas inundáveis de várzea e igapó
São florestas que sofrem inundação durante parte do ano. Ocorrem nas margens do rio Xingu, sendo mais comuns na zona do baixo curso. Em alguns trechos, aparecem formações com as palmeiras jauari (Astrocaryumjauari), açaí (Euterpe oleracea) e caranã (Mauritiella armata) nas zonas mais rebaixadas do relevo. No entanto, não chegam a configurar um elemento de paisagem. São florestas localizadas na planície de inundação, com desnível variando entre 4 e 8 m nos picos anuais de enchente e vazante. Os solos são geralmente de origem hidromórfica, do grupo glei húmico; são de drenagem deficiente e incoporam considerável teor de matéria orgânica e nutrientes anualmente.
O dossel deste tipo de floresta aluvial é menos compacto e fechado do que na tipologia densa, localizando-se a uma altura de aproximadamente 20 m. As espécies mais comuns neste estrato são o tarumã (Vitex triflora, Violaceae), ipê da várzea (Tabebuia barbata, Bignoniaceae), xixuá (Maytenus sp., Celastraceae) e o ipê (Macrolobium acaciaefolium, Caesalpiniaceae). O sub-bosque desta floresta é limpo, com pouca regeneração das espécies do dossel. Provavelmente, a mortalidade produzida pela inundação selecione poucas mudas, reduzindo drasticamente o número de indivíduos jovens. Algumas poucas espécies são deste estrato, geralmente de porte reduzido e talvez com adaptação ecofisiológica para conviver com submersão. Foram registradas neste estrato Oxandra riedelinana (Annonaceae) e Ticorea longifolia (Rutaceae).
Dentre as árvores emergentes, tem-se o açacu (Hura crepitans, Euphorbiaceae), a piranheira (Piranhea trifoliolata, Euphorbiaceae), a abiurana da várzea (Pouteria glomerata, Sapotaceae) e acapurana (Campsiandra laurifolia, Caesalpiniaceae).
A floresta aluvial, regionalmente conhecida como várzea, possui uma diversidade pouco inferior à floresta de terra firme em função das condições hidrológicas onde a lâmina d'água varia consideravelmente. Em geral, cerca de 120 a 150 espécies de árvores são encontradas neste ambiente, distribuídas em famílias botânicas como Caesalpiniaceae e Euphorbiaceae. A abundância de indivíduos entre as espécies é melhor distribuída do que nas florestas densas de terras baixas.
Floresta ombrófila aberta com palmeira - Floresta de terra firme com palmeiras
Este tipo de floresta é freqüente na parte sudeste e sul do Pará; sudoeste do Maranhão, seguindo o arco sul amazônico, que inclui o norte do Mato Grosso, Rondônia, chegando até o Acre.
A fisionomia é caracterizada pela presença de grandes palmeiras com altura de até 30 m. Entre as palmeiras, destacam-se espécies como o babaçu (Attalea speciosa), inajá (Attalea maripa), paxiúba (Socratea exorhiza), bacaba (Oenocarpus bacaba) e, em menor escala, o patauá (O. bataua).
Na região de Belo Monte, as florestas com palmeiras estão concentradas na porção sul da Volta Grande, na margem direita do rio Xingu; pode ser encontrada também no centro da área de estudo, na margem esquerda.
Essa tipologia apresenta a segunda maior área dentro da cena estudada, desenvolvendo-se geralmente sobre latossolos profundos e podzólicos. Estas florestas abertas têm uma maior flamabilidade, sendo freqüentes incêndios em áreas onde foi feita exploração madeireira, mesmo que ela seja seletiva.
O conjunto das características de história natural desta tipologia é similar àquela encontrada na floresta densa, uma vez que ela compartilha grande parte de suas espécies. No entanto, as espécies das florestas abertas, sejam elas com palmeiras ou com cipós, apresentam maior grau de cadufolia, podendo chegar, em alguns casos, a 10% do total das espécies. Podem ser consideradas florestas semideciduais, associadas ao déficit hídrico existente nos meses menos chuvosos, em outubro e novembro. A proporção de espécies cujos propágulos são dispersos pelo vento também aumenta nas florestas abertas, onde a presença de dossel descontínuo deve facilitar o mecanismo de anemocoria, ao contrário do que acontece nas florestas densas.
As espécies arbóreas mais abundantes, dominantes e freqüentes, associadas às florestas de palmeiras, na região de Belo Monte, são: acapu (Vouacapoua americana, Caesalpini aceae), melancieira (Alexa grandiflora, Fab aceae) mão-de-gato (Helicostylis tomentosa, Moraceae), castanheira-do-pará (Bertholletia excelsa, Lecythidaceae), cariperana (Licania egleri, Chrysobalan aceae), abiurana (Pouteria lasiocarpa, Sapotaceae), envira preta (Guatteria poeppigiana, Annonaceae), ingá-pereba (Inga alba, Mimosaceae), aroeira (Astronium lecointei, Anacardiaceae), paricá (Schyzolobium amazonicum, Caesalpiniaceae), araracanga (Aspidosperma araracanga, Apocynaceae), dentre outras.
O dossel da floresta é descontínuo, dominado pela palmeira babaçu (Attalea speciosa), de grande porte e que pode atingir 30 m de altura, permitindo a passagem de luz solar total; proliferam lianas (cipós), arbustos e ervas no sub-bosque. As emergentes podem atingir alturas consideráveis, em torno de 50 m.
A diversidade das florestas abertas com palmeiras é também considerável, sendo que boa parte das espécies são compartilhadas com as florestas densas. Em termos gerais, essas florestas suportam riqueza de espécies que varia entre 140 e 180 espécies arbóreas, cipós lenhosos e palmeiras por hectare.
Floresta ombrófila aberta com cipós e palmeiras - Floresta de terra firme com cipós e palmeiras
Na região de Belo Monte, as florestas ombrófilas abertas com cipós e palmeiras constituem uma tipologia característica, embora estejam dentro da zona onde a paisagem é mais fragmentada pela ação humana. Na área estudada, essas florestas abertas localizam-se em terreno submontanhoso, com altitude variando entre 150 e 300 m. A paisagem destas florestas é marcada pela disposição espaçada das árvores, o que favorece a colonização por lianas e palmeiras. Esses grupos vegetais dependem de luz para se desenvolver plenamente.
Os cipós proliferam e, em alguns casos, podem matar algumas árvores. É comum observar as 'torres de cipós', constituídas pela colonização das lianas em árvores mortas. Entre os cipós, destacam-se o rabo de arara (Acacia multipinnata, Mimosaceae), escada de jabuti (Bauhinia guianensis, Caesalpiniaceae), Leucocalantha aromatica (Bignoniaceae) e cipó-abuta (Abutagrandifolia). Nas áreas mais baixas dos vales, aparecem as palmeiras açaí (Euterpe oleracea, Arecaceae) e paxiúba (Socratea exorhiza, Arecaceae).
Dentro da área estudada, as florestas abertas com cipós e palmeiras localizam-se na margem esquerda do rio Xingu, à parte nordeste da área trabalhada, ao leste da sede do município. É a tipologia mais impactada por diversos usos da terra, como agricultura familiar e pastagens, sendo considerável também o percentual de áreas de capoeiras abandonadas.
As florestas abertas crescem sobre solos rasos com afloramentos rochosos, com pouca à mediana retenção hídrica, o que produz considerável caducifolia para os padrões de florestas ombrófilas amazônicas (em torno de 10% também), denotando certo grau de sazonalidade. Segundo Veloso et al. (1991), as florestas de cipós estariam em estágio anticlímax, produzido por flutuações climáticas dos períodos mais secos no passado, que provavelmente não voltaram ao estágio clímax por restrições edáficas atuais.
O dossel das florestas abertas densas permite a passagem de cerca do dobro de luz solar em relação à floresta densa, proporcionando o franco desenvolvimento e colonização de espécies lucíferas, tanto arbóreas, como arbustivas, herbáceas e lianescentes.
As principais espécies florestais associadas às florestas abertas com cipós e palmeiras são a castanheira (Bertholletia excelsa, Lecythidaceae), melancieira (Alexa grandiflora, Fabaceae), pau de remo (Chimarrhisturbinata, Rubiaceae), cacau do mato (Theobroma speciosum, Sterculiaceae), ipês amarelo e roxo (respectivamente, Tabebuia serratifolia e T impetiginosa, Bignoniaceae), acapu (Vouacapoua americana, Caesalpiniaceae), muiracatiaras (Astronium gracile e A. lecointei, Anacardiaceae), geniparana (Gustavia augusta, Lecythidaceae), tatajuba (Bagassa guianensis, Moraceae), dentre outras.
A diversidade das florestas abertas, inventariadas na área estudada, é menor do que aquela registrada para as florestas densas. A riqueza de espécies variou entre 119 e 128 por hectare. As espécies raras, consideradas aquelas com densidade menor ou igual a 1 indivíduo/hectare, são bem representadas, algo em torno de 40% a 50% do total de espécies.
RESULTADOS E DISCUSSÃO
O sresultados doravante apresentados foram subitemizados em florística, fitossociologia, estrutura e etnobotânica e particularizados nas diversas fitofisionomias. Uma outra premissa adotada refere-se à estratificação diamétrica do ecossistema florestal: considera-se floresta propriamente dita aquela formação vegetal cujos espécimes amostrados têm DAP ≥ 10 cm, e sub-bosque o estrato cujos indivíduos estão abaixo deste limite e acima de 5 cm de DAP.
Na região de Belo Monte, como frisado an teriormente, foram analisadas as vegetações referentes a quatro fitofisionomias da floresta ombrófila: floresta ombrófila densa, floresta ombrófila aluvial, floresta ombrófila aberta com palmeira e floresta ombrófila aberta com cipós e palmeiras, sendo também analisada a floresta ombrófila densa com exploração seletiva de madeira.
FLORÍSTICA
Foram registrados, em todas as fitofisionomias, 13.790 indivíduos compreendendo 662 espécies distribuídas em 65 famílias. A relação de todas as espécies registradas é apresentada no Apêndice 1, com a respectiva forma de vida, número total de indivíduos amostrados e o respectivo uso/utilidade, por fitofisionomia analisada. O número total de indivíduos, espécies e famílias, nas diversas classes diamétricas analisadas, é apresentado na Tabela 4.

Famílias botânicas
Considerando-se as fitofisionomias com vegetação primária, observa-se que a abundância, diversidade e número de famílias dos indivíduos (DAP ≥ 10 cm) atingem os maiores valores médios na floresta ombrófila densa, floresta ombrófila aberta com cipó e palmeira e floresta ombrófila aluvial, respectivamente; já a floresta ombrófila densa com exploração seletiva de madeira apresenta valores próximos à floresta ombrófila com cipó e palmeira (Tabela 5).
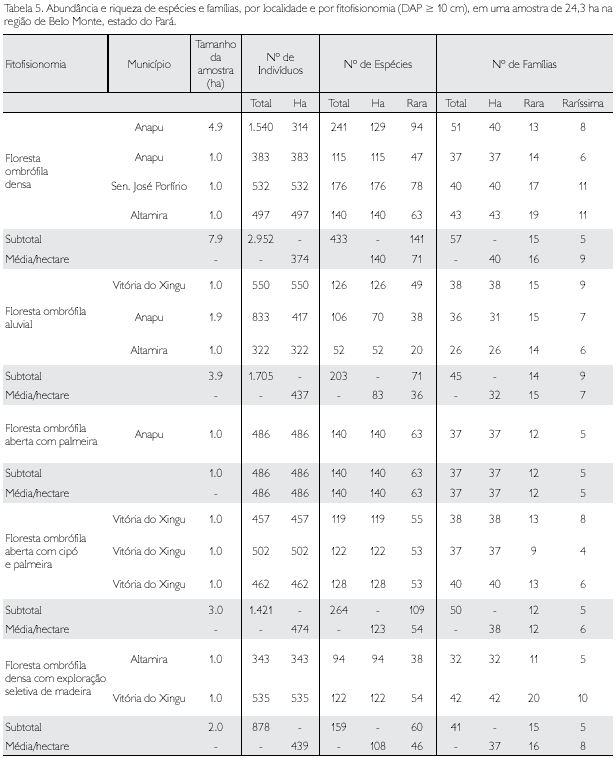
As estimativas médias das fitofisionomias retratam com certa fidelidade a realidade de cada ecossistema florestal. Para gerar estimativas mais próximas das médias verdadeiras, deve-se procurar uniformizar a amostragem no que tange ao método, tamanho, forma e número de parcelas, além de estabelecer uma intensidade de amostragem adequada à área física de cada fitofisionomia estudada. Quando se analisa, por exemplo, as espécies raras, sabe-se que, em média, na Amazônia Oriental há cerca de 450 árvores/hectare e algo como 130 a 140 espécies (DAP ≥ 10 cm), sendo que 1/3 dessas ocorrem com apenas um indivíduo para cada hectare amostrado.
Todavia, ao se aumentar o número de parcelas, percebe-se que essa 'raridade' vai decaindo, em decorrência da área de dispersão das espécies estar se ampliando. Outra implicação de não haver um número razoável de unidades amostrais recai sobre um sub ou super dimensionamento de variáveis como número de árvores, volume de madeira, área basal e biomassa.
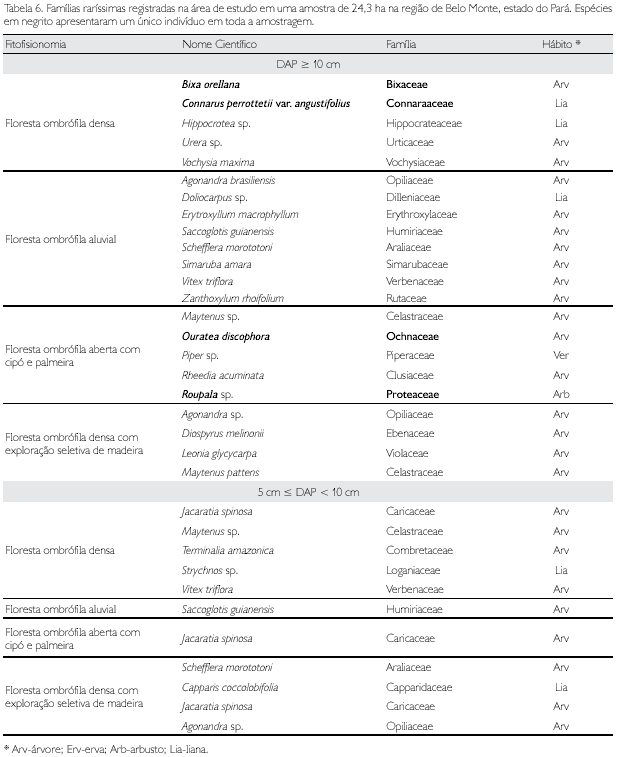
Considera-se família 'rara' aquela com uma única espécie e 'raríssima' seria aquela com uma única espécie e um único indivíduo. Curiosamente, percebe-se que, para todas as fitofisionomias analisadas, existem cinco famílias raríssimas, exceto para a floresta ombrófila aluvial, que tem nove famílias (Tabela 5). As espécies dessas famílias são muito importantes num programa de conservação, sobretudo aquelas em negrito, pois em toda a amostragem suas espécies apresentaram um único indivíduo (Tabela 6).
A maior riqueza, considerando-se a amostragem total da vegetação, foi de Mimosaceae, com 55 espécies, seguindo-se Sapotaceae (52), Fabaceae (41), Caesalpiniaceae (39) e Moraceae (32); no intervalo entre dez e 30 espécies foram amostradas 15 famílias, e com menos de dez e mais de uma espécie existem outras 29 famílias; com apenas uma espécie amostrou-se 16 famílias (Apêndice 1).
Para a abundância, considerando-se também a amostragem total, o maior valor foi da família Arecaceae (1.480 indivíduos), seguindo-se Caesalpiniaceae (1.259) e Mimosaceae (1.078); com mais de 100 e menos de 1.000 indivíduos, observam-se 22 famílias; e entre 100 e dois indivíduos outras 36 famílias - Bixaceae, Connaraceae, Ochnaceae e Proteaceae, como já referido, são as famílias que apresentaram um único indivíduo e, conseqüentemente, também uma única espécie (Apêndice 1).
Considerando-se a família Leguminosae, composta por suas três subfamílias, como proposta por Cronquist (1981), observa-se que, de fato, é a mais rica (135 espécies), a mais abundante (2.782 indivíduos) e, conseqüentemente, a de maior diversidade.
Espécies
A variação na riqueza e composição de espécies em regiões tropicais tem sido atribuída a diversos fatores, tais como tipos de solo, relevo, disponibilidade de energia, variação na precipitação, temperatura, altitude e distância (Wright et al., 1993). Condit et al. (2002) relatam que a alta riqueza nas florestas tropicais, particularmente na Amazônia, tem sido amplamente documentada. Contudo, os fatores bióticos e abióticos que influenciam a composição de espécies têm sido pouco estudados.
As espécies de maior abundância na amostragem total foram a palmeira açaí (Euterpe oleraceae), com 675 indivíduos; a melancieira (Alexa grandiflora), com 348; geniparana (Gustavia augusta), com 346; acapu (Vouacapoua americana), com 287; cupuí (Theobroma speciosum), com 25; trapiarana (Leonia sp.), com 230; e muruci d'anta (Molia luscens), com 225 indivíduos. Entre 100 e 200 indivíduos, amostraram-se 24 espécies, sendo três de palmeiras-pati (Syagrus sp.), com 192; mumbaca (Astrocaryum mumbaca), com 175; babaçu (Attalea speciosa), com 164; e uma de cipó (cipó escada - Bauhinia sp.), com 114; as demais são espécies arbóreas. Com menos de 100 e mais de dois indivíduos, amostrou-se 486 espécies e com apenas um indivíduo foram registradas 144 espécies (Apêndice 1).
Analisando-se a capacidade das espécies amostradas em se adaptarem nas diversas fitofisionomias estudadas (plasticidade), observou-se que 289 espécies ocorreram em apenas uma fitofisionomia, em contraste a outras 23, que ocorreram em todas as fitofisionomias amostradas. Ou seja, 44% do total das espécies têm plasticidade restrita (mínima ou grau 1 - presente em apenas um dos cinco ecossistemas analisados), enquanto apenas 3,5% têm plasticidade ampla (máxima ou grau 5 - presente nos cinco ambientes). Com distribuição em duas, três e quatro fitofisionomias, registrou-se, respectivamente, 164, 115 e 71 espécies (Apêndice 1).
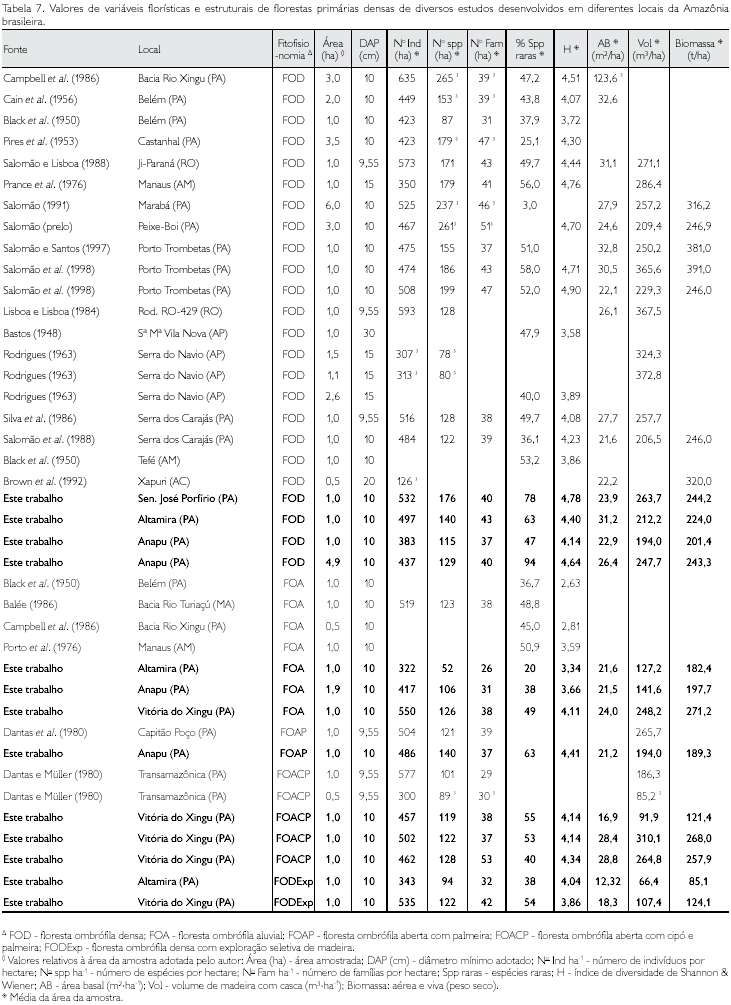
Segundo Ducke e Black (1954), é um fato estranhável que na hiléia (Amazônia) a longitude desempenhe um papel muito mais importante que a latitude na composição florística: a diferença das floras é muito mais acentuada entre Belém e Santarém que entre Belém e Caiena (Guiana Francesa), embora, no último caso, a distância seja maior. Várias observações acusam número maior de espécies para o centro e noroeste da Amazônia que para as partes orientais e ocidentais da região (Tabela 7).
Diversidade
Para a análise de diversidade, foram utilizados os índices de Shannon & Wiener (Ricklefs, 1979), de eqüabilidade (Pielou, 1975) e o índice de Simpson (Simpson, 1949). A variação da diversidade em diversas escalas é um dos conceitos mais difundidos para explicar quais fatores controlam a diversidade em comunidades ecológicas (Pitman et al., 1999). Diversos estudos têm sido publicados relacionados à variação de riqueza e composição de espécie em diferentes distâncias geográficas, mostrando que estas variações são resultantes de fatores climáticos (Ter Steege et al., 2002). Contudo, poucos estudos testaram a mudança na composição de espécies em escala local (Ferreira et al., prelo).
O índice de Shannon & Wiener, por ser sensível ao número de espécies raras, é diretamente influenciado pelo conjunto de espécies raras da comunidade (Gomes et al., 2004). Uma crítica que se faz a esse índice reside no fato de que o cálculo da diversidade, baseado na abundância relativa, não permite uma resposta ecológica plausível de análise após o seu cálculo (Magurran, 1988).
A eqüabilidade é a relação entre o índice de Shannon & Wiener encontrado e o valor máximo possível para o mesmo número de espécies, expresso quando todas as espécies apresentam o mesmo número de indivíduos, ou seja, é a propriedade de uma comunidade que diz respeito à uniformidade de distribuição de espécies ou suas abundâncias relativas. Eqüabilidade máxima significa uniformidade máxima, e eqüabilidade mínima, quando há uma espécie dominante. Por exemplo, uma amostra contendo 200 indivíduos em dez espécies, com 20 indivíduos de cada espécie, tem mais eqüidade que uma amostra de mesmo tamanho e número de espécies, em que uma das espécies tem 100 indivíduos e as outras nove espécies compartilham os outros 100 indivíduos.O índice de Simpson (D) expressa a probabilidade de quaisquer dois indivíduos tomados aleatoriamente na amostra pertencerem à mesma espécie (Simpson, 1949); por exemplo, D = 1 para uma monocultura e D = 0 para uma comunidade diversa composta de indivíduos únicos de cada espécie. Tal índice é muito influenciado pelas espécies mais abundantes e é insensível àquelas espécies que aparecem com apenas um indivíduo na amostra. Via de regra, como dito anteriormente, na Amazônia Oriental (Tabela 7), tem-se observado que geralmente em uma amostra de 1 ha - DAP ≥ 10 cm - há cerca de 450 indivíduos distribuídos em 130-140 espécies, sendo que 1/3 dessas espécies ocorrem com um único indivíduo. Em síntese, este índice é pouco influenciado pela riqueza de espécies, pesando mais a abundância das espécies com maior freqüência.
Magurran (1988) definiu o índice de Simpson como a probabilidade de dois indivíduos, tomados ao acaso, pertencerem a diferentes espécies: a expressão matemática apresentada é justamente a probabilidade de se tomar dois indivíduos da mesma espécie. O complementar (1 - D) é muitas vezes usado como índice de diversidade, já que D é um valor de dominância na comunidade - esta expressão é a probabilidade a que se refere Magurran (l.c).
Os índices de diversidade (H' e 1-D), dominância (D) e eqüabilidade (J) entre as fitofisionomias analisadas mostraram que as comunidades são altamente diversas (Tabela 8). O índice de diversidade de espécies de Shannon & Wiener (H'), de acordo com Margalef (1968), normalmente apresenta valores entre 1,5 a 3,5, raramente ultrapassando 4,5 para logaritmo neperiano; observa-se que, na floresta ombrófila densa aqui analisada, ficou o valor próximo a 5,3, variando nos demais entre 4,2 e 4,8. O índice de diversidade de Simpson (1-D) é de praticamente 1 (máximo) na floresta ombrófila densa e inferior a 0,99 nas demais fitofisionomias.
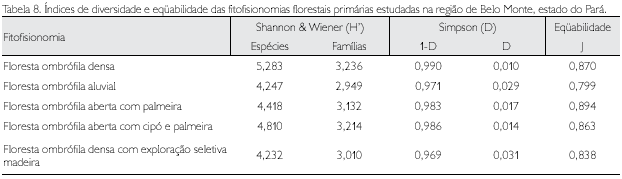
O índice de dominânica de Simpson (D) indicou que nas fitofisionomias não houve dominância de quaisquer espécies ou foi muito baixa, ficando entre 1 (floresta ombrófila densa) e 2,9% (floresta ombrófila aluvial) a probabilidade de se amostrar dois indivíduos ao acaso que pertençam à mesma espécie. Na floresta com exploração seletiva de madeira, como esperado, a probabilidade foi maior (Tabela 8).
A eqüabilidade também foi alta, indicando que os valores de diversidade de Shannon & Wiener ficaram próximos ao máximo esperado para o número de espécies amostradas, o que demonstrou que a grande maioria das espécies contribuiu com n úmeros de indivíduos bem próximos nas fitofisionomias estudadas (Tabela 8).
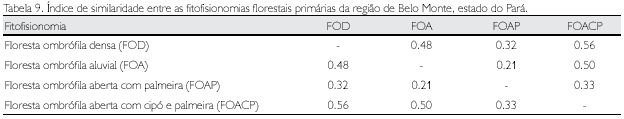
Similaridade
Existem vários critérios para quantificar a similaridade ou dissimilaridade entre comunidades biológicas de duas áreas. As propriedades do índice têm importância prática, já que seu valor é estimado a partir de amostras. Um bom índice deve ter alta precisão na estimativa de similaridade entre comunidades e baixo potencial de viezes.
Segundo Ferreira et al. (prelo), diversos autores relatam que a distância geográfica é considerada uma das variáveis que mais influenciam a distribuição das espécies em regiões tropicais (Ruololainen; Tuomisto, 2002). Condit et al. (2002), comparando inventários florísticos entre Panamá, Peru e Equador, encontraram uma correlação negativa entre a similaridade florística das parcelas em função da distância geográfica. Scudeller et al. (2001) também demonstraram que existe uma correlação negativa entre similaridade florística e a distância geográfica em uma floresta ombrófila densa da Mata Atlântica no Brasil, enquanto Carneiro e Valeriano (2003) encontraram uma forte correlação negativa entre a distância geográfica e a similaridade florística, comparando comunidades de plantas em relação à variação da altitude e distância geográfica na Mata Atlântica no Brasil.
Oliveira e Mori (1999) relatam uma similaridade florística variando de 30% a 35% em parcelas de amostradas em uma floresta ombrófila densa, próximo a Manaus, Amazonas. Ferreira e Prance (1998) encontraram um padrão semelhante em uma floresta ombrófila densa no Parque Nacional do Jaú/lbama, estado do Amazonas. Almeida et al. (2003) encontraram uma similaridade variando entre 31% e 37%, em parcelas de floresta ombrófila densa no Parque Ambiental de Gunma, em Belém, Pará, na Amazônia Oriental.
Essa baixa similaridade de espécies entre parcelas de vegetação na Amazônia está relacionada ao baixo número de espécies compartilhadas entre essas, podendo estar associado a fatores como a grande proporção de espécies raras e alta variabilidade de habitats entre as parcelas (Almeida et al., 2003), a biologia floral e ao estágio sucessional das espécies (Ferreira et al., prelo).
Para análise da similaridade florística entre as fitofisionomias estudadas, calculou-se o índice qualitativo de Sorènsen (1943), que representa a probabilidade de se sortear uma espécie comum às duas amostragens, considerando que elas têm o dobro de chance de serem amostradas do que as espécies que aparecem em apenas uma das amostragens.
As maiores similaridades de espécies observadas ocorreram entre a floresta ombrófila aberta com cipó e palmeira, a floresta ombrófila densa (0,56) e a floresta ombrófila aluvial (0,50). Já a maior dissimilaridade (0,21) foi observada entre a floresta ombrófila aberta com palmeira e a floresta ombrófila aluvial (Tabela 9). A floresta ombrófila densa, como era esperado, por apresentar maior riqueza de espécies, é a que mais fortemente se assemelha qualitativamente com as demais fitofisionomias estudadas.
FITOSSOCIOLOGIA
No Xingu, Campbell et al. (1986) investigaram três hectares (3.000m x 10m) de mata de terra firme e meio hectare (500m x 10m) de várzea, utilizando amostragem em transectos e critérios de inclusão de DAP ≥ 10 cm. Esta metodologia foi adotada porque os autores queriam 'amostrar o maior número de espécies por unidade de área'. Na mata de terra firme, foram encontradas 1.420 indíviduos, 39 famílias e 265 espécies, sendo Cenostigma macrophyllum Tul. e Orbignya sp. as espécies mais importantes (maior IVI); as famílias mais importantes foram Leguminosae, Palmae (Arecaceae), Lecythidaceae, Moraceae e Bombacaceae. Em cada hectare foram encontradas 393, 460 e 567 árvores e 133, 118 e 162 espécies, respectivamente. Apenas quatro espécies (15%) foram registradas nos três hectares e a similaridade variou entre 2,6 a 3,35 (índice de Jaccard) para cada hectare. Os autores concluíram que a diversidade da floresta era ocasionada pelo grande número de espécies raras que contribuíam com muito pouco do total do lVl da comunidade e que a Amazônia é um mosaico de diferentes tipos de florestas, não podendo se extrapolar os dados de riqueza de espécies de pequenas amostragens para áreas mais amplas (Oliveira, 1997).
Na área de influência da Usina Hidrelétrica (UHE) de Kararaô, no Xingu, Cardenas (1987), em uma área de 15 ha de floresta de terra firme e 10 ha de floresta densa ciliar, registrou 54 famílias que foram relacionadas de acordo com o valor de importância de família (VlF). As famílias Caesalpiniaceae (176 indivíduos), Burseraceae (153) e Moraceae (111) foram as três mais importantes da terra firme; nas floretas ciliares as famílias mais importantes foram Meliaceae (123 indivíduos), Lecythidaceae (116) e Annonaceae (108). O total de espécies registradas nesse inventário foi de 280 e de 176 gêneros. Na floresta de terra firme, a maior abundância foi de Tetragastris altíssima (118 indivíduos), Voucapoua americana (105) e Theobroma sylvestre (67); na floresta densa ciliar foi de Carapa guianensis (79 indivíduos), Conceveiba sp. (74) e Virola surinamensis (73).
Poucos estudos foram desenvolvidos na região de Belo Monte e para um melhor entendimento da fitofisionomia dessa exuberante região, apresentar-se-á a seguir a análise fitossociológica para cada uma das paisagens de vegetação identificadas e amostradas em campo.
Floresta ombrófila densa
Foram identificadas 57 famílias nos quatro municípios de abrangência desta fitofisionomia (Tabela 5). Catorze indivíduos (12 cipós e dois arbóreos) ficaram indeterminados neste nível taxonômico devido à indisponibilidade de material botânico para identificação (Tabela 10). Leguminosae, se considerada como uma única família botânica, teria nítida dominância sobre as demais: lVl = 26,3% contra 8,3% da segunda colocada (Lecythidaceae). Mimosaceae, Sapotaceae, Caesalpiniaceae e Moraceae foram as famílias com maior riqueza, apresentando, respectivamemte, 43, 42, 32 e 21 espécies (31,9% do total).
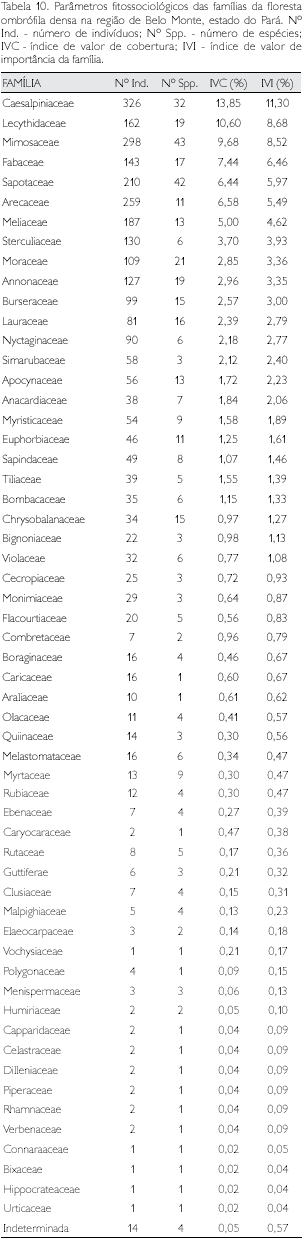
Entre 10 e 20 espécies, obtiveram-se 10 famílias e, entre duas e nove espécies, observaram-se 28 famílias, praticamente a metade do número de famílias. Com uma única espécie, verificou-se um total de 15 famílias, consideradas raras neste estudo. Cinco famílias (Bixaceae, Connaraceae, Hippocrateaceae, Urticaceae e Vochysiaceae) apresentaram um único indivíduo e, conseqüentemente, uma única espécie, sendo consideradas rarríssimas (Tabelas 5 e 10). As sete famílias que apresentaram os maiores valores de lVl corresponderam a mais de 51% do total desta variável, respondendo, também, por 40,7% da diversidade total e 53,7% da abundância total nesta fitofisionomia (Tabela 10).
Num universo de 2.952 indivíduos amostrados com DAP ≥ 10 cm, foram identificadas 433 espécies distribuídas em 57 famílias, cujos parâmetros fitossociológicos são apresentados no Apêndice 2. As dez espécies que apresentaram maiores valores de IVI respondem por mais de 20% do total desta variável, enquanto o número de indivíduos corresponde a 22,5% do total; 143 (ou 32,3% do total) apresentaram um único indivíduo (Apêndice 2); tal fato tem implicação direta no baixo índice de similaridade dentre as fitorregiões analisadas (Tabela 9), bem como num programa de conservação, pois essas espécies são as mais vulneráveis.
Inga alba, Sterculia pruriens, Tachigalia myrmecophila e Vouacapoua americana, entre as de maior IVI, acham-se presentes em todas as fitorregiões analisadas, enquanto 139 espécies (21%), do total de 662 amostradas em todas as fitorregiões, são exclusivas da floresta ombrófila densa (Apêndice 1).
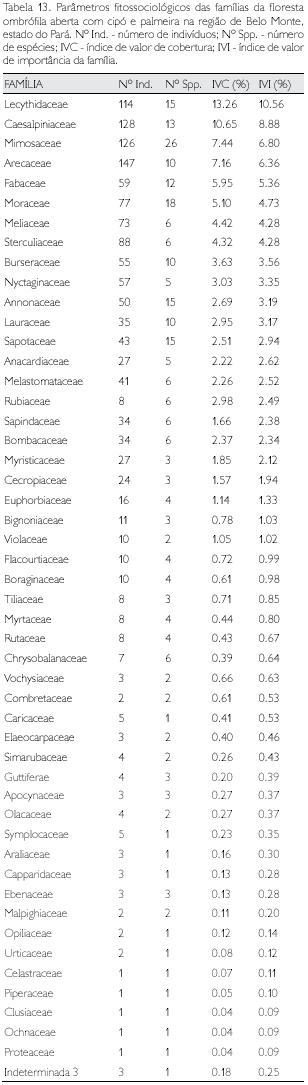
Floresta ombrófila aluvial
Foram identificadas 45 famílias nos três municípios (Tabela 5) de abrangência da amostragem (3,9 ha) entre os indivíduos com DAP ≥ 10 cm. Oito famílias, equivalentes a 18% do total - Caesalpiniaceae (16 espécies), Fabaceae (15), Sapotaceae (14), Mimosaceae (12), Moraceae (12), Burseraceae (10), Lecythidaceae (11) e Chrysobalanaceae (11) - apresentaram praticamente 50% do total das espécies; com número de espécies entre duas e nove, amostrou-se 24 famílias (Tabela 13). Apresentando uma única espécie, registrou-se um total de 13 famílias, consideradas raras neste estudo, e aquelas nove famílias que apresentaram um único indivíduo (Tabela 11).
As famílias que apresentaram maior abundância foram Caesalpiniaceae (288 indivíduos) e Fabaceae (220), entre 100 e 200 indivíduos registrou-se quatro famílias e com mais de um espécime e menos de 100 outras 32 famílias; nove famílias foram consideradas raríssimas nesta fitofisionomia. O somatório do IVI das cinco famílias que apresentaram os maiores índices corresponde a mais da metade do IVI total, 50,8% (Apêndice 3). Leguminosae lato sensu teria nítida dominância sobre as demais: IVI = 35,9% contra 8,3% da segunda colocada (Lecythidaceae).
Num universo de 1.705 indivíduos amostrados (DAP ≥ 10 cm), foram identificadas 203 espécies distribuídas em 45 famílias (Tabela 5), cujos parâmetros fitossociológicos são apresentados no Apêndice 3. As dez espécies que apresentaram maiores valores de IVI respondem por mais de 41% do total desta variável, enquanto o número de indivíduos corresponde a 43,2% do total de indivíduos. Setenta e uma espécies (35%) apresentaram um único indivíduo (Apêndice 3).
Quarenta e três espécies são exclusivas desta fitofisionomia, destacando-se Macrolobium sp. (com 139 indivíduos), Discocarpum spruceanum (62), Leonia psycarpa (38), Capsiandra laurifolia, Eugenia lambertiana, Neoxythece cuspidata, Vismia sp. (cada uma com dez indivíduos); apenas Alexa grandiflora (melancieira) ocorreu em todas as outras fitofisionomias (Apêndice 1).
Floresta ombrófila aberta com palmeira
Foram identificadas 37 famílias no único município de abrangência da amostra dos indivíduos com DAP ≥ 10 cm nesta fitofisionomia (Tabela 5). Sete famílias equivaleram a 18,9% do total, Mimosaceae (14 espécies), Sapotaceae (13), Annonaceae, Burseraceae e Moraceae (cada uma com 9), Lauraceae (8) e Lecythidaceae (7) apresentaram praticamente 50% do total da riqueza de espécies (Tabela 16). Com uma única espécie verificou-se um total de 12 famílias consideradas raras e aquelas cinco famílias que apresentaram um único indivíduo e, conseqüentemente, uma única espécie (Tabela 12).
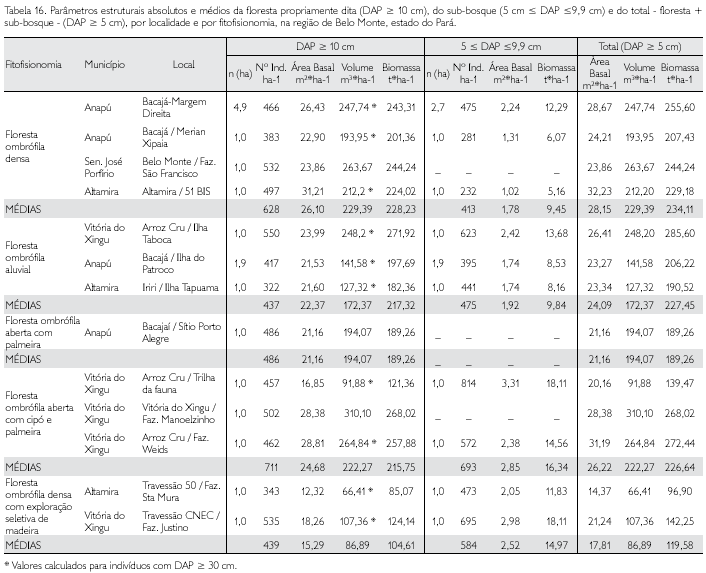
As famílias que apresentaram maior abundância foram Moraceae (55 indivíduos), Mimosaceae (45) e Caesalpiniaceae (40); com abundância entre dez e 40 indivíduos registraram-se 13 famílias, e com mais de um e menos de dez espécimes outras 16 famílias; cinco famílias foram consideradas raríssimas nesta fitofisionomia. O somatório do IVI das sete famílias que apresentaram os maiores índices corresponde a mais da metade do IVI total - 52,5% (Tabela 12). Leguminosae lato sensu teve nítida dominância sobre as demais: IVI = 25,7% contra 9,1% da segunda colocada (Moraceae) (Tabela 12).
Nessa amostragem, foram registrados 486 indivíduos (DAP ≥ 10 cm), totalizando 140 espécies distribuídas em 37 famílias (Tabela 5), cujos parâmetros fitossociológicos são apresentados no Apêndice 4. As dez espécies que apresentaram maiores valores de IVI respondem por pouco mais de 33% do total desta variável, enquanto o número de indivíduos corresponde a 32,5% do total (Apêndice 4).
Seis espécies entre as dez de maior IVI (Vouacapoua americana, Alexa grandiflora, Inga alba, Helicostylis tomentosa, Pouteria lasiocarpa e Guatteria poeppigiana) ocorreram em todas as outras fitofisionomias (Apêndice 1).
Mais da metade das espécies (73) apresentou um único indivíduo (Apêndice 4). Como já frisado anteriormente, esse fato tem implicação direta num programa de conservação, pois essas espécies seriam as mais vulneráveis; 29 espécies são exclusivas desta fitofisionomia, destacando-se Matayba arborescens (10 indivíduos), Virola elongata (8) e Lacunaria crenata (7) (Apêndice 1).
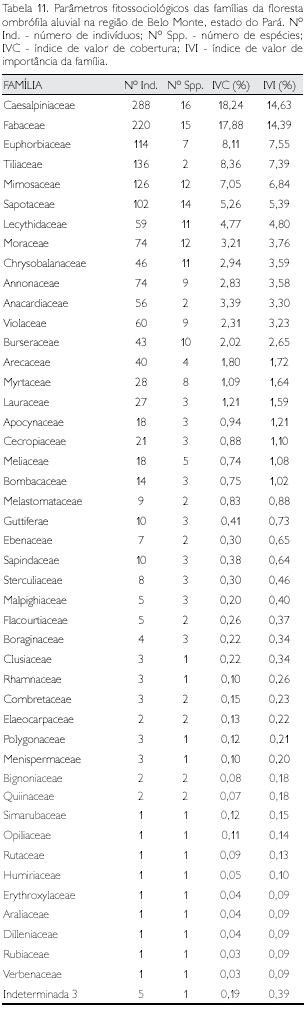
Floresta ombrófila aberta com cipó e palmeira
Foram identificadas 50 famílias nos três municípios (Tabela 5) de abrangência da amostragem dos indivíduos com DAP ≥ 10 cm nesta fitofisionomia. As 10 famílias com maior lVl (equivalente a 51,3% do total) apresentaram mais da metade do total das espécies; Mimosaceae (26 espécies) foi a família com maior diversidade, seguindo-se Moraceae (18), Lecythidaceae, Annonaceae e Sapotaceae (cada uma com 15 espécies); entre 10 e 13 espécies têm-se cinco famílias e entre duas e seis exatamente metade (27) do total de famílias; com uma única espécie verificou-se 13 famílias consideradas raras (Tabela 13) e, também, cinco famílias que apresentaram um único indivíduo e, conseqüentemente, uma única espécie, consideradas raríssimas (Tabela 6).
As famílias que apresentaram maior abundância foram Arecaceae (147 espécimes), Caesalpiniaceae (128), Mimosaceae (126) e Lecythidaceae (114); entre 10 e 100 indivíduos foram amostradas 21 famílias e com mais de um espécime e menos de 10 outras 21 famílias (Tabela 13). O lVlF acumulado das nove famílias que apresentaram os maiores valores corresponde a mais da metade do lVlF total - 51,3%.
No total de 1.421 indivíduos (DAP ≥ 10 cm) amostrados em 3 ha, foram identificadas 264 espécies distribuídas em 50 famílias (Tabela 5), cujos parâmetros fitossociológicos são apresentados no Apêndice 5. As dez espécies que apresentaram maiores valores de lVl respondem por 27,8% do total desta variável, enquanto o número de indivíduos corresponde a 345 (24,3% do total); 71 espécies (35,0%) apresentaram um único indivíduo (Apêndice 5).
Sete espécies entre as dez de maior lVl ocorrem em todas as outras demais fitofisionomias (Bertholletia excelsa, Alexa grandiflora, Vouacapoua americana, Theobroma speciosum, Euterpe oleracea, Inga alba e Tachigali mymercophila) . Quarenta e seis espécies são exclusivas dessa fitofisionomia, destacando-se Dimorphandra sp. (com 8 indivíduos), Coussapoua sp. (6), Pseudolmedia murure e Symplocos guianensis (cada uma com cinco espécimes) (Tabela 21).
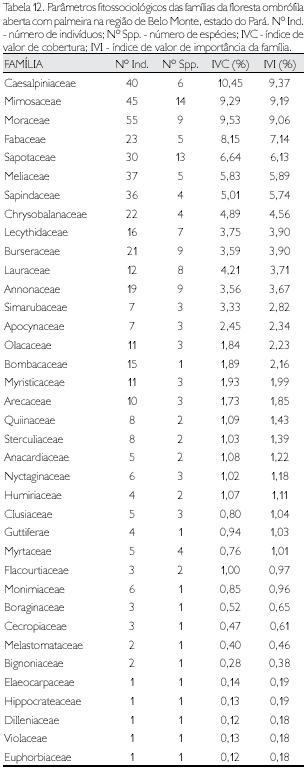
Floresta ombrófila densa com exploração seletiva de madeira
Foram identificadas 41 famílias nos dois municípios (Tabela 5) de abrangência da amostragem (2 ha) dos indivíduos com DAP ≥ 10 cm nessa fitofisionomia antrópica. Dez famílias, equivalente a 24,4% do total, apresentaram mais da metade do total das espécies; com uma única espécie verificou-se um total de 15 famílias consideradas raras neste estudo, enquanto outras cinco famílias apresentaram um único indivíduo (Tabela 14) e, conseqüentemente, uma única espécie, sendo consideradas raríssimas (Tabela 6).
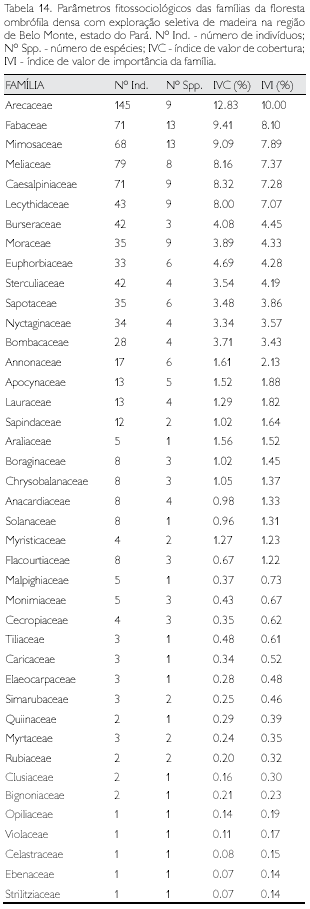
As famílias que apresentaram maior abundância foram Arecaceae (145 indivíduos), Meliaceae (79), Fabaceae e Caesalpiniaceae (com 71 espécimes cada) e Mimosaceae (68); entre 50 e 10 indivíduos amostrou-se 12 famílias e com mais de um espécime e menos de dez outras 20 famílias. O somatório do IVI das oito famílias que apresentaram os maiores índices corresponde a mais da metade do IVI total - 51,9% (Tabela 14); cinco famílias foram consi deradas raríssimas nesta fitofisionomia (Tabela 6).
Num total de 878 indivíduos (DAP ≥ 10 cm) amostrados em 2 ha, foram identificadas 159 espécies distribuídas em 41 famílias (Tabela 4), cujos parâmetros fitossociológicos são apresentados no Apêndice 6; as dez espécies que apresentaram maiores valores de IVI respondem por 32% do total desta variável; o número de indivíduos destas corresponde a 331 (37,7% do total). Sessenta espécies (37,7%) apresentaram um único indivíduo (Apêndice 6).
Nesta fitofisionomia, foram retiradas todas as espécies de valor madeireiro que apresentavam DAP ≥ 40 cm (Apêndice 1), conseqüentemente, a riqueza florística decaiu, assim como as variáveis estruturais área basal, volume de madeira e biomassa. Quatro espécies (Alexa grandiflora, Bertholletia excelsa, Theobroma speciosum e Vouacapoua americana) ocorreram em todas as demais fitofisionomias (Apêndice 1).
ESTRUTURA DENDROMÉTRICA
Estrutura diamétrica
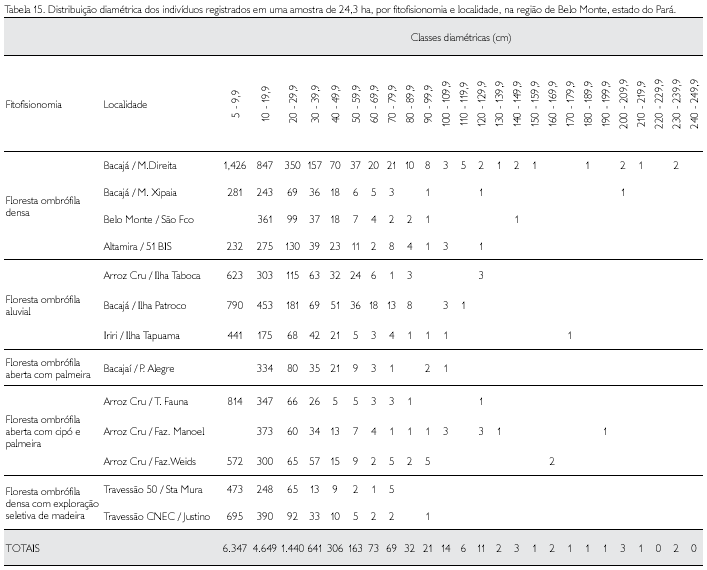
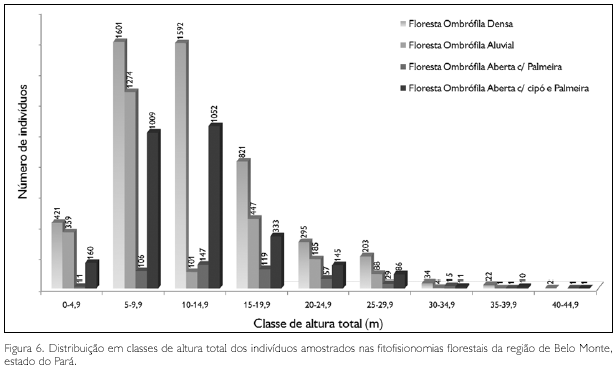
A distribuição diamétrica de todos os 13.789 indivíduos, com limite de DAP ≥ 5 cm, por fitofisionomia, é apresentada na Tabela 15. Os espécimes que apresentaram os maiores diâmetros foram Bertholletia excelsa, castanheira (234,9 cm, 232,1 cm, 213,3 cm e 206,9 cm), uma tanimbuca (Terminalia amazonica) com 203,7 cm, e um exemplar de amarelão (Apuleia mollaris) com 202,1 cm. Mais de 46% dos indivíduos apresentaram DAP entre 5 cm e 10 cm; praticamente 34% têm diâmetros entre 10 cm e 20 cm, ou seja, mais de 80% dos indivíduos amostrados têm diâmetro igual ou inferior a 20 cm. O diâmetro médio da amostra foi de 15,2 cm ± 14,4 cm; acima dessa média tem-se apenas 17% do total de indivíduos.
A distribuição diamétrica dos indivíduos da floresta ombrófila densa (FOD) é apresentada na Figura 3. Todos aqueles espécimes mencionados no parágrafo anterior, que são os maiores em diâmetro entre todas as amostragens, pertencem a essa fitofisionomia, que apresentou 27 árvores com DAP ≥ 100 cm (Tabela 15).
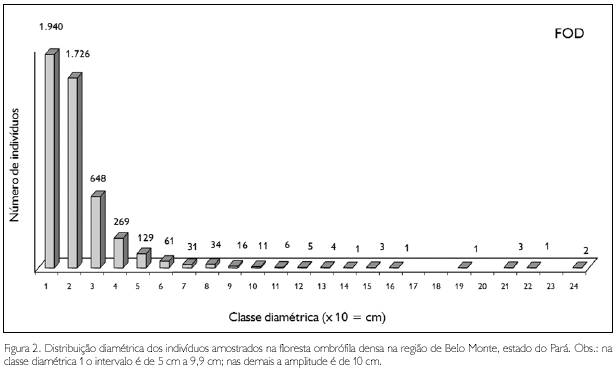
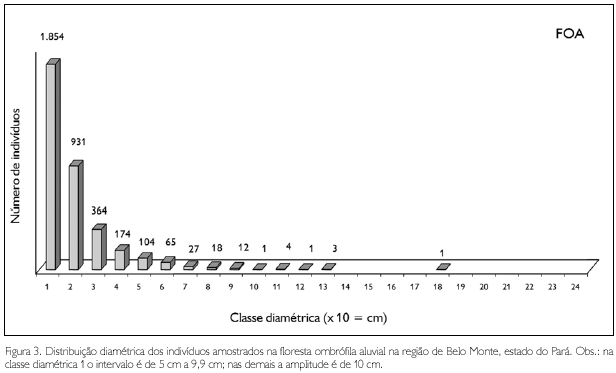
Na floresta ombrófila aluvial (FOA), os maiores diâmetros observados foram em um matamatá branco (Eschweilera coriacea) com 170,3 cm, sapucaia (Lecythis pisonis) com 127,3 cm, um tauari (Couratari guianensis) com 124,1 cm, e numa mirauba (Mouriri myrtiflora) com 121 cm; apenas nove espécimes apresentaram diâmetro superior a 100 cm (Figura 4).
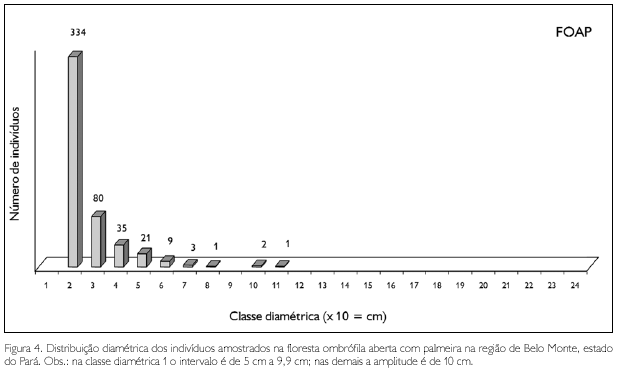
A floresta ombrófila aberta com palmeira apresentou uma única árvore com diâmetro superior a 100 cm: marupa amarelo (Simaba paraensis), com 104,4 cm. Imediatamente após, observou-se um exemplar de melancieira (Alexa grandiflora) e outro de itauba (Mezilaurus itauba), com, respectivamente, 94,8 cm e 93,4 cm; todas as demais apresentaram DAP < 70 cm (Figura 5).
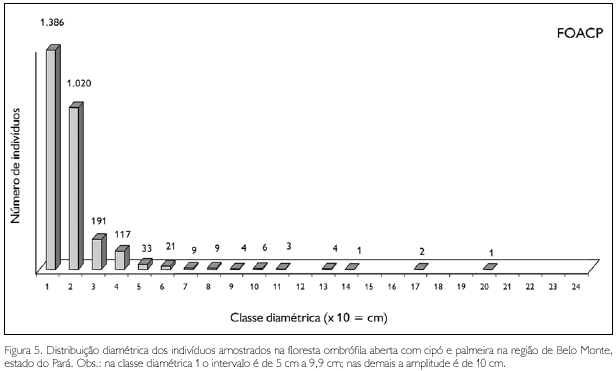
Dez árvores apresentaram DAP ≥ 100 cm na floresta ombrófila aberta com cipó e palmeira, destacando-se: pau de remo (Chimarris turbinata) com 190 cm, castanheira e um tachi preto (Tachigalia mymercophila), ambas com 162,3 cm; outros seis espécimes foram de castanheira, com diâmetros variando entre 127,3 cm e 108 cm, e a restante de maior diâmetro foi outro 'pau de remo', com 101,7 cm (Figura 6).
Estrutura de altura
Nas florestas da região do Xingu, geralmente, as grandes árvores apresentam altura total inferior às de outras regiões da Amazônia, como as da região do rio Trombetas, de Curuá-Una, de Marabá e de Caxiuanã. Nestes locais, as maiores árvores alcançam de 50 m a 60 m. Em todas as amostragens efetuadas no Xingu, a maior altura estimada foi de 40 m em espécimes de amarelão (Apuleia molaris), tauari (Couratari guianensis), tachi preto e castanheira.
A altura total média de todas as árvores nas florestas primárias da região estudada foi de 11,2 m ± 5,6 m; mais da metade das árvores (50,4%) estão abaixo deste limite médio (Figura 7).
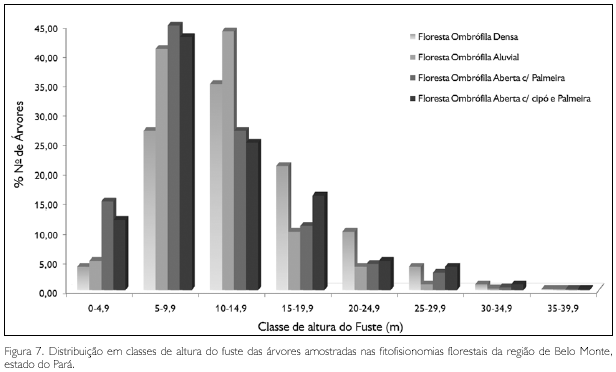
A altura do fuste atingiu o valor máximo em um espécime de castanheira e outro de melancieira, ambos com 35 m; acima de 30 m apenas um outro exemplar de morototó (Schefflera morototoni), com 31 m. A distribuição das árvores em classes de fuste, ou tronco (Figura 8), evidencia que 58,7% apresentaram altura do fuste inferior à média, que foi de 11,3 m ± 5,5 m.
Área basal
A área basal é um importante índice de aferimento das formações florestais. Para isto, convencionou-se arbitrariamente admitir o limite mínimo de 10 cm de diâmetro para a inclusão dos troncos na amostragem. Este limite otimiza o trabalho e é aproximadamente o limite de diâmetro dos espécimes que atingem o dossel ('teto') da mata, além de existir mais dados práticos coletados para comparações. Assim, se for dito que uma floresta tem 40 m2*ha-1 de área basal, sabe-se tratar de uma floresta muito exuberante, com muitas árvores grossas (diâmetros acima de 1 e 2 m), como a da região de Trombetas; um campo com 0,5 m2*ha-1 é um campo limpo; já as matas de cipó da região de Marabá e Itacaiunas ficam entre 18 m2*ha-1 e 22 m2*ha-1 (Pires, 1981).
Nas florestas primárias da região do Xingu, o maior valor de área basal, entre todas as fitofisionomias analisadas, foi aquele sob o domínio da floresta ombrófila densa, na localidade denominada 51 Bis, em Altamira, que totalizou 31,21 m2*ha-1 e o menor foi de 16,85 m2*ha-1, na localidade de Arroz Cru, em Vitória do Xingu, sob o domínio da floresta ombrófila com cipó e palmeira (Tabela 16).
Os valores médios da área basal nas diversas fitofisionomias foram: 26,1 m2*ha-1, 24,68 m2*ha-1, 22,37 m2*ha-1, 21,16 m2*ha-1 e 15,29 m2*ha-1, respectivamente, na floresta ombrófila densa, floresta ombrófila aberta com cipó e palmeira, floresta ombrófila aluvial, floresta ombrófila aberta com palmeira e floresta ombrófila densa com exploração seletiva de madeira (Tabela 16).
Volumetria
O volume médio de madeira estimado para a floresta ombrófila densa foi de 229,39 m3*ha-1; para a floresta ombrófila aluvial de 172,37 m3*ha-1; para a floresta ombrófila aberta com cipó e palmeira de 222,27 m3*ha-1; para a floresta ombrófila com palmeira de 194,07 m3*ha-1; e para a floresta explorada foi de 86,89 m3*ha-1 (Tabela 16).
Na floresta ombrófila com cipó e palmeira, tanto o volume quanto a área basal e a biomassa apresentaram valores altos para as localidades da Fazenda Manoelzinho e da Weids (Tabela 16). Tal fato, muito incomum, é explicado porque na primeira localidade ocorreram dois exemplares de pau de remo com 190 cm e 101,7 cm, cinco de castanheiras com DAP variando de 131 cm a 108 cm e um de tanimbuca com 100; ou seja, oito árvores com DAP superior a 100 cm que, em conjunto, respondem por 143,8 m3*ha-1 do volume (ou 46,4% do total), 10,2 m2*ha-1 da área basal (ou 35,9% do total) e 116,6 t*ha-1 de biomassa (ou 43,5% do total). Deve-se considerar, também, que na segunda localidade ocorreram uma castanheira e um tachi preto, ambos com 162,3 cm de diâmetro (somente estas duas árvores possibilitaram um incremento de 25,2%, 14,4% e 16,8% do total do volume, área basal e biomassa, respectivamente).
A floresta ombrófila densa é a que apresentou o maior potencial madeireiro, o que explica a floresta explorada desta fitofisionomia apresentar um volume médio de apenas 86,89 m3*ha-1, permitindo projetar que exista algo como 142,5 m3*ha-1 de madeira comercializável nestas florestas (Tabela 16).
A floresta ombrófila aberta com cipó e palmeira da região do Xingu apresenta maior volume de madeira que aquelas estudadas por Dantas e Müller (1980), na Transamazônica. Já a floresta ombrófila densa do Xingu apresenta volume de madeira semelhante às demais localidades da Amazônia (Tabela 7).
Biomassa
A biomassa aérea viva, por fitofisionomia e para cada localidade amostrada, é apresentada na Tabela 16, separadamente para todos os indivíduos com DAP ≥ 10 cm e 5 cm ≤ DAP < 10 cm (sub-bosque) e, também, totalizada independentemente para cada classe diamétrica (estrato).
A estimativa da biomassa aérea viva das árvores com DAP ≥ 5 cm, da floresta ombrófila densa foi de 234,11 t*ha-1 (toneladas, em peso seco, por hectare), na floresta ombrófila aluvial foi de 227,45 t*ha-1, na floresta ombrófila com cipó e palmeira foi de 226,64 t*ha-1 e na floresta explorada de 119,58 t*ha-1; na floresta ombrófila aberta com palmeira não foram mensurados indivíduos entre 5 cm e 9,9 cm, daí se dispor apenas da estimativa de biomassa para DAP ≥ 10 cm, que foi de 189,26 t*ha-1 (Tabela 16).
A quantificação da biomassa das florestas neotropicais apresenta uma grande amplitude entre as estimativas de diversos estudos, variando de 185 t*ha-1, na Colômbia, a 406 t*ha-1, no Brasil (Salomão et al., 1988). Essas discrepâncias são resultados de diversas incertezas, mas, sobretudo, dois fatores são os mais importantes: a extrapolação ou interpretação errônea da tipologia florestal e a inclusão (total ou parcial) ou não das diversas frações da biomassa total nas estimativas apresentadas (Tabela 7).
Comumente, as estimativas de biomassa da floresta tropical primária, em vários estudos, referem-se à fração majoritária da biomassa total, que é a biomassa aérea viva (troncos, galhos e folhas) das árvores com, geralmente, DAP ≥ 10 cm. Todavia, como já foi dito, a biomassa total apresenta diversos componentes que devem ser quantificados para que as estimativas geradas cada vez mais se aproximem da média verdadeira. Em alguns estudos, a biomassa morta corresponde a aproximadamente 14% da biomassa total, cabendo à biomassa viva o percentual restante. A biomassa aérea viva corresponde a pouco menos de 70% da total - em muitos estudos, essa fração é considerada como sendo a biomassa total - a biomassa viva subterrânea equivale a aproximadamente 20% da biomassa total ou a 25% da biomassa viva (Salomão et al., 1988).
A biomassa aérea calculada nesse estudo é uma aproximação subdimensionada por não se dispor de dados referentes a todos os componentes desta, como troncos mortos e em pé com DAP < 10 cm e, também, da biomassa total das plantas com DAP < 5 cm.
A análise da compartimentação dessa biomassa na floresta primária permite inferir que praticamente metade (c.a. 46%) das árvores amostradas por hectare responde por apenas 5,2% da biomassa total - esses são os indivíduos formadores do sub-bosque da floresta e têm o DAP compreendido entre 5 cm e 10 cm. As árvores com DAP ≥ 50 cm (2,9% do total de árvores) respondem por 52,3% da biomassa total de todas as fitofisionomias - praticamente a mesma biomassa que as 97,1% das árvores restantes (Tabela 16). Esse dado é interessante quando se analisa o estoque de carbono e, conseqüentemente, as emissões de carbono de áreas de extração seletiva de madeira de valor econômico, pois, muito raramente, árvores com menos de 50 cm são abatidas para comercialização. A primeira vista, essas áreas podem ser confundidas como sendo de florestas primárias, na interpretação e análise de imagens geradas por satélite, e superdimensionar as áreas de floresta primária que apresentam biomassa minimamente superior ao dobro destas.
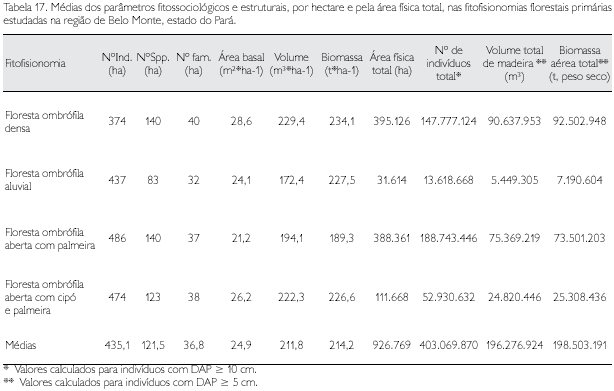
Calculou-se um total de 92,68 km2 de cobertura florestal primária distribuída nas quatro fitofisionomias florestais analisadas para a região de Belo Monte, gerando, em termos de abundância, uma estimativa de 403.069.870 árvores com DAP ≥ 10 cm, um volume de madeira da ordem de 196.276.924 m3 e uma estimativa da biomassa aérea viva em torno de 198.503.191 t (peso seco), para toda a região.
A árvore média da região do Xingu apresenta os seguintes valores estruturais médios: DAP de 15,2 cm ± 14,4 cm, altura total de 11,2 m ± 5,6 m, altura do fuste de 11,3 m ± 5,5 m, área seccional de 0,042 ± 0,007 m2, volume de madeira com casca de 0,368 ± 0,040 m3 e biomassa aérea viva de 0,388 ± 0,081 t.
ESPÉCIES ARBÓREAS RELEVANTES
Foram identificadas algumas espécies florestais de grande interesse para os programas de conservação da flora. Foi observado, no município de Senador José Porfírio, a presença de uma espécie arbórea praticamente extinta nas áreas onde é possível a penetração humana: trata-se do pau cravo (Dicypellium caryophyllatum), uma espécie de Lauraceae muito cobiçada no passado pela indústria perfumista, que praticamente a dizimou das matas. Outra espécie de díficil ocorrência é a Sagotia brachysepala (arataciú), uma euforbiácea de dispersão muito restrita. A castanheira (B. excelsa) e a seringueira (Hevea brasiliensis) são espécies protegidas por instrumentos legais que não permitem seu corte, sendo que ambas têm ocorrência natural na área de estudo; estas espécies devem ter programas específicos de preservação/conservação in situ e, talvez, também ex situ.
Existem algumas espécies que apresentam característi cas m u i to p ecu li ares, com o Acacia multipinnata, que é um cipó muito agressivo ao ponto de, onde se desenvolve, levar a vegetação arbórea a um declínio acentuado, pois sua biomassa foliar impede as árvores do dossel de receber luz; em contrapartida, existem outras que são excelentes fixadoras de nitrogênio, como Chamaecrista diphylla - ambas espécies ocorreram na amostragem.
Uma outra preocupação que deve ser observada nos programas de conservação refere-se às fitofisionomias registradas. Verifica-se que as áreas originalmente cobertas pela floresta ombrófila densa e a floresta ombrófila aberta com palmeira estão dispersas majoritariamente na área estudada; em contrapartida, a floresta ombrófila aluvial com apenas pouco mais de 31.000 ha deve, necessariamente, ter porções significativas sob proteção legal (Tabela 17).
ETNOBOTÂNICA
Salomão et al. (1995) avaliaram 108 espécies arbóreas amazônicas quanto aos usos não-madeireiros (alimento humano e da fauna, fibras, medicinais, produtoras de gomas, celulose, valor ornamental, tintura, subtância ven en osa, con stru ções rú sti cas) e as resp ectivas distribuições diamétricas nos ecossistemas primários da floresta amazônica.
Shanley e Rosa (2005) avaliaram o uso não-madeireiro (alimento humano e da fauna, materiais de construção, remédios, tecnologia, comércio e outros usos como práticas espirituais/magias) de 153 espécies arbóreas inventariadas ao longo do rio Capim, no município de Ipixuna, no estado do Pará.
As 662 espécies identificadas e registradas no presente trabalho foram classificadas quanto à utilidade/uso atual (Apêndice 1). Para algumas destas espécies, não foi possível identificar o uso ou utilidade atual, seja por desconhecimento ou falta de material bibliográfico disponível (ausência de estudos específicos). Sinteticamente, a distribuição do número de espécies com a respectiva utilidade é apresentada na Tabela 18.
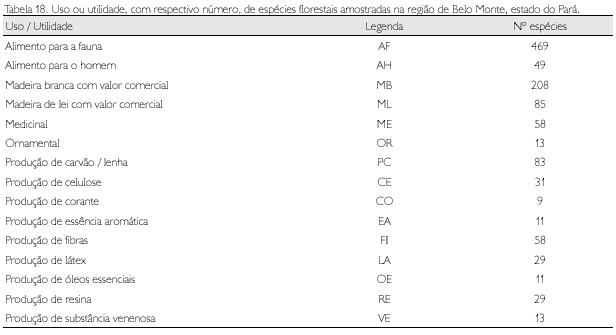
Algu m as con si d erações acerca desses u sos devem ser feitas: (1) quaisquer plantas cujos frutos são comestíveis para o homem também o serão pelos animais, todavia, a recíproca não é verdadeira; (2) várias espécies apresentam mais de um uso ou utilidade, por exemplo, a maioria das Lauraceae serve tanto de alimento para fauna (fruto) como são produtoras de essências aromáticas; (3) algumas espécies produzem substâncias venenosas/tóxicas para o homem, animais ou para ambos.
Se considerado que todas as árvores servem pelo menos para fazer sombra e produzir lenha, todas as 662 espécies arbóreas amostradas têm utilidade. Todavia, considerando-se o valor da madeira, observa-se que 44,3% delas têm valor comercial. Alternativamente, com uso e/ou utilidade não madeireira, há 55,7% das espécies (Figura 8). Salomão et al. (1995) descreveram uma breve caracterização ecológica-econômica de 108 espécies arbóreas da Amazônia brasileira, espécies que também são encontradas nas florestas do Xingu.
Doze espécies (Bertholletia excelsa, Bixa orellana, Cedrela odorata, Euterpe oleracea, Hymenaea courbaril, Inga alba, Inga cinammomea, Jacaratia spinosa, Lecythis pisonis, Richardella macrophylla, Symphonia globulifera e Virola surinamensis) apresentaram o maior número de usos/utilidades (quatro); outras 102, 317 e 171 espécies apresentaram, respectivamente, três, dois e u m u sos; p ara 60 esp éci es n ão foi possível a caracterização etnobotânica, pelo desconhecimento ou pela identificação taxonômica incompleta da espécie (Apêndice 1).
CONCLUSÃO
A riqueza das florestas ombrófilas amazônicas é uma das mais elevadas da região neotropical, variando de 115 a 241 espécies de árvores com DAP ≥ 10 cm. A distribuição da abundância de indivíduos nessas florestas registra um elevado número de espécies localmente raras, c.a. 1/3 do total de espécies apresentando um indivíduo por hectare, em contraposição a um outro grupo restrito de espécies com elevado número de indivíduos.
A floresta ombrófila densa é uma das mais representativas da Amazônia e, por conseguinte, na área de estudo. Nos inventários realizados na região do rio Bacajá e próximo à vila de Belo Monte, a abundância variou de 314 a 532 árvores por hectare, quando se considera somente o segmento com DAP ≥ 10 cm.
A floresta ombrófila aluvial, regionalmente conhecida como várzea, possui uma diversidade razoável e as condições hidrológicas, como a lâmina d'água, variam consideravelmente. Em geral, cerca de 120 a 150 espécies arbóreas são encontradas neste ambiente, distribuídas em famílias botânicas representativas, como Caesalpiniaceae e Euphorbiaceae; a abundância entre as espécies é melhor distribuída do que nas florestas densas de terras baixas.
A riqueza das florestas ombrófilas abertas com palmeiras é também considerável, sendo que boa parte das espécies são compartilhadas com as florestas densas. Em termos gerais, essas florestas suportam uma riqueza de espécies que varia entre 140 a 180 espécies arbóreas, cipós lenhosos e palmeiras por hectare.
A riqueza das florestas ombrófilas com cipós e palmeiras é menor do que aquela registrada para as florestas densas. A riqueza de espécies variou entre 119 e 128 por hectare. As espécies raras totalizaram 40% a 50% do total de espécies.
Comparando-se as fitofisionomias estudadas entre os ecossistemas primários, pode-se concluir que aquela com maior riqueza é a floresta ombrófila densa, seguindo-se a floresta ombrófila aberta com cipó e palmeira, a floresta ombrófila aluvial e a floresta ombrófila aberta com palmeira.
Com relação às espécies raras, a floresta ombrófila densa apresentou o maior número, em contraposição à floresta ombrófila aberta com palmeira, que apresentou o menor número; a floresta ombrófila aluvial foi a que apresentou o maior número de espécies raríssimas.
Caesalpiniaceae foi a família que apresentou o maior IVI e IVC em todas as fitofisionomias estudadas, à exceção da floresta ombrófila aberta com cipó e palmeira. Alexa grandiflora e Voucapoua americana foram as espécies de maiores índices em todas as fitofisionomias estudadas, exceto na floresta ombrófila aluvial, com Pterocarpus amazonicus e Molia luscens se sobressaindo.
Foram identificadas algumas espécies florestais de grande interesse para os programas de conservação da flora. Foi verificada, no município de Senador José Porfírio, a presença de uma espécie arbórea praticamente extinta nas áreas onde é possível a penetração humana; trata-se do pau cravo (Dicypellium caryophyllatum), uma Lauraceae muito cobiçada pela indústria perfumista, que praticamente a dizimou das matas. Outra espécie de díficil ocorrência é a arataciú (Sagotia brachysepala), uma euforbiácea de dispersão muito restrita.
AGRADECIMENTOS
Às Centrais Elétricas do Norte do Brasil - Eletronorte que, através do convênio MCT-MPEG/ELETRONORTE, 2002, financiou esta pesquisa.
REFERÊNCIAS
AB'SABER, A. N. The paleoclimate and paleoecology of Brazilian Amazonia. In: WHITMORE, T. C.; PRANCE, G. T. (Eds.). Biological diversification in the tropics. New York: Columbia University Press, 1982. 714 p. p. 41-59.
ABSY, M. L. Quaternary palynological studies in the amazon basin. In: WHITMORE, T. C.; PRANCE, G. T. (Eds.) Biological diversification in the tropics. New York: Columbia University Press, 1982. 714 p. p. 67-73.
ALMEIDA, S. S.; LISBOA, P. L. B.; SILVA, A. S. Diversidade florística de uma comunidade arbórea na estação científica "Ferreira Penna" em Caxiuanã (Pará). Bol. Mus. Para. Emílio goeldi, Belém, v. 9, n. 1, p. 93-128, 1993.
ALMEIDA, S. S.; AMARAL, D. D.; SILVA, A. S. M.; THALES, M. Inventário florístico e análise fitossociológica dos ambientes do Parque Ecológico de gumma, Município de Santa Bárbara, PA. Belém: MPEG/MCT-JICA, 2003. 188 p. Relatório Técnico.
AYRES, J. M. As matas de várzea do Mamirauá: médio Solimões. 2. ed. Brasília: CNPq/Sociedade Civil Mamirauá, 1995.
BALÉE, W. Análise preliminar de inventário florestal e etnobotânica Ka'apor (Maranhão). Bol. Mus. Para. Emílio goeldi, ser. Bot., Belém, v. 2, n. 2, p. 141-67, 1986.
BASTOS, A. M. As matas de Santa Maria do Vila Nova, Território do Amapá. An. Bras. de Economia Florestal, v. 1, p. 121-8, 1948.
BLACK, G. A.; DOBZANSKY, T; PAVAN, C. Some attenpts no estimate species diversity and population density of trees in Amazonian Forest. Bot. geog, v. 111, p. 413-25, 1950.
BROWN, S.; GILLESPIE, A. J. R.; LUGO, A. Biomass estimation methods for tropical forests with applications to forest inventory data. Forest Science, v. 35, n. 4, p. 881-902, 1989.
BROWN, I. F; NEPSTAD, D. C.; PIRES, I. O.; LUZ, L. M.; ALECHAMDRE, A. S. Carbon storage and Land-use in Extractive Reserves, Acre, Brazil. Environmental Conservation, v. 19, n. 4, p. 307-15, 1992.
CAIN, S. A.; CASTRO, G. M. A.; PIRES, J. M.; SILVA, N. T. Application of some phytosociological techniques to brasilian rain forest. An. J. Bot., v. 43, p. 911-41, 1956.
CAMPBELL, D. G.; DALY, D. C.; PRANCE, G. T; MACIEL, U. M. Quantitative ecological inventory of terra firme and várzea tropical forest on the Rio Xingu, brasilian Amazon. Brittonia, v. 38, n. 4, p. 369-93, 1986.
CAMPBELL, D. G. P; HAMMOND, H. D. Floristic inventory of tropical countries. New York: New York Botanical Garden, 1989. 545 p.
CARDENAS, J. D. R. Levantamento e análise da fitomassa da UHE de Kararaô, Rio Xingu. Manaus: INPA, 1987. 182 p. Relatório Técnico.
CARNEIRO, J. S.; VALERIANO, D. M. Padrão espacial da diversidade beta da Mata Atlântica -Uma análise da distribuição da biodiversidade em um banco de dados geográficos. In: SBSR, São Paulo, 11, 2003. Anais... São Paulo: INPE, 2003. p. 629-36.
CONDIT, R.; PITMAN, N.; LEIGHT, JR. E. G.; CHAVE, J.; TERBORGH, J.; FOSTER, R. B.; NUNEZ, P. V.; AGUILAR, S.; VALENCIA, R.; VILLA, G.; MULLER-LANDAU, C.; LOSOS, E.; HUBBEL, S. P. Beta diversity in tropical trees. Science, v. 295, p. 666-8, 2002.
COTTAM, G.; CURTIS, J. T. The use of distance mensures in phytosociological sampling. Ecology, v. 37, n. 3, p. 451-60, 1956.
CRONQUIST, A. An integrated system of classification of flowering plants: The New York Botanical Garden. New York: Columbia University Press, 1981. 1262 p.
DALY D.; PRANCE, G. T. Brazilian Amazon. In: CAMPBELL, D. G. P. Harmond. Floristic inventory of tropical countries. New York: New York Botanical Garden, 1989. 545 p. p. 402-424.
DANTAS, M.; MÜLLER, N. R. M. Estudos fito-ecológicos do trópico úmido brasileiro I -Aspectos fitosociológicos de mata de terra roxa na região de Altamira. In: CONGRESSO NACIONAL DE BOTÂNICA, 30, São Paulo, 1980. Anais... São Paulo, 1980. p. 205-18.
DANTAS, M.; RODRIGUES, J. A.; MÜLLER, N. R. M. Estudos fito-ecológicos do trópico úmido brasileiro: aspectos fitosociológicos de mata sobre latossolo em Capitão Poço, PA. Bol. Pesq. Embrapa/ Cpatu, Belém, v. 9, p. 1-19, 1980.
DUCKE, A.; BLACK, G. A. Notas sobre a fitogeografia da Amazônia braileira. Bol. Téc. do Inst. Agron. Norte, Belém, v. 29, p. 1-48, 1954.
FALESI, I. C. Os solos da rodovia Transamazônica. Bol. Técn. IPEAN, v. 55, p. 1-196, 1972.
FERREIRA, L. V.; PRANCE, G. T Structure and species richness of low diversity floodplain forest on the rio Tapajós, Eastern Amazonian. Biodiversity and Conservation, v. 7, n. 5, p. 585-96, 1998.
FERREIRA, L. V; SALOMÃO, R. P.; PEREIRA, J. L. G.; ROSA, N. A.; BARLOW, J.; MORAIS, K. A. C. Padrão espacial da similaridade de espécies arbóreas em função da distância em floresta ombrófila densa na Amazônia setentrional: implicações para a conservação da biodiversidade, (no prelo).
FORSTER, M. Strukturanalyses eines tropischen regenwaldes in Kolunbien. Allg. Forst. -u. J.-Ztg., v. 144, n. 1, p. 1-8, 1973.
GOMES, B. Z.; MARTINS, F. R.; TAMASHIRO, J. Y. Estrutura do cerradão e da transição entre cerradão e floresta paludicola num fragmento da Internacional Paper do Brasil Ltda., em Brotas, SP. Revista Brasileira de Botânica, v. 27, n. 2, p. 249-62, 2004.
HAFFER, J. General aspects o euge theory. In: Whitmore, T. C.; Prance, G. T. (Eds.). Biological diversification in the tropics. New York: Columbia University Press, 1982. 714 p. p. 6-26.
HUECK, K. As florestas da América do Sul. São Paulo: USP 1966.
LISBOA, P L. B.; LISBOA, R. C. L. Inventário florestal em Rondônia. I - Rodovia Presidente Médici - Costa Marques, Km 90. In: CONGRESSO NACIONAL DE BOTÂNICA, 35, Manaus, 1984. Anais... Manaus, 1984.
MARGALEF, R. Perspectives in Ecological Theory. Chicago: University of Chicago Press, 1968. 112 p.
MAGURRAN, A. E. Ecological diversity and its measurement. New Jersey: Princeton University Press, 1988.
MIRANDA, E. E; MIRANDA, J. R. de; SANTOS, E. P. F. dos. Efeitos ecológicos das barragens do Xingu: uma avaliação preliminar. In: LEINAD, Ayer; SANTOS, O.; ANDRADE, L. M. M. de (Orgs.). As hidrelétricas do xingu e os povos indígenas. São Paulo: Comissão Pró-Índio de S. Paulo, 1988. 196 p.
MUSEU PARAENSE EMÍLIO GOELDI. Diagnóstico da vegetação e da flora da área de estudo da Usina Hidrelétrica de Belo Monte, Estado do Pará. Belém: Convênio MCT-MPEG/ELETRONORTE, 2002. Relatório Técnico. 171 p.
MUELLER-DOMBOIS, D.; ELLEMBERG, H. Aims and methods of vegetation ecology. New York: J. Willey and Sous, 1974. p. 45-135.
OLIVEIRA, A. A. Diversidade, estrutura e dinâmica do componente arbóreo de uma floresta de terra firme de Manaus, Amazonas, 1997. 155 p. Tese (Doutorado) - Universidade de São Paulo, Instituto de Biociências, São Paulo, 1997.
OLIVEIRA, A. A.; MORI, S. A. A central Amazonian terra firme forest. I. High tree species richness on poor soils. Biodiversity and Conservation, v. 8, p. 1219-44, 1999.
PIELOU, E. C. Ecological diversity. New York: J. Wiley, 1975. 403 p.
PIRES, J. M. Tipos de vegetação da Amazônia. Belém: MPEG, 1973. p. 179-202. (Publicações Avulsas do Museu Paraense Emílio Goeldi, 20).
PIRES, J. M. O conceito de floresta tropical. CVRD-revista, v. 2, n. 5, p. 17-20, 1981.
PIRES, J. M. The Amazonian Forest. In: SIOLI, H. The Amazon: limnology and landscape ecology of a mighty tropical river and its basin, Dordrecht: W. Junck publishers, 1984. 763 p. p. 581-602.
PIRES, J. M.; PRANCE, G. T. The vegetation types of the Brazilian Amazon. In: PRANCE, G.T.; LOVEJOY, T. E. Key environments: Amazonia. Oxford: Pergamon Press, 1985. 442 p. p. 109-145.
PIRES, J. M.; DOBZANSKY, T.; BLACK, G. A. An estimate of the number ofspecies of trees in an Amazonian Forest Community, Bot. Gaz., v. 114, n. 4, p. 467-77, 1953.
PITMAN, N. C. A.; TERBORG, J.; SILMAN, M. R.; V. NUNEZ, P. Tree species distribution in an upper amazonian forest. Ecology, v. 80, n. 8, p. 2651-2661, 1999.
PORTO, M. L.; LONGHI, H. M.; CITADINI, V.; RAMOS, R. F.; MARIATH, J. E. A. Levantamento fitossociológico em área de "mata-de-baixio" na Estação Experimental de Silvicultura Tropical, INPA, Manaus. Acta Amazônica, Manaus, v. 6, n. 3, p. 301-318, 1976.
PRANCE, G. T.; RODRIGUES, W. A.; SILVA, M. F. Inventário florestal de um hectare de mata de terra firme, Km 30 da estrada Manaus - Itacoatiara. Acta Amazônica, Manaus, v. 6, n. 1, p. 9-35, 1976.
PRANCE, G. T.; LOVEJOY, T. E. Key environments: Amazonia. Oxford: Pergamon Press, 1985. 442 p.
PRANCE, G.T. Forest refuges: evidences from woody angiosperms. In: WHITMORE, T. C.; PRANCE, G. T. (Eds.). Biological diversification in the tropics. New York: Columbia University Press, 1979. 714 p. p. 137-6.
PROJETO RADAMBRASIL. Folha SA.22. Belém: DNPM, 1974. Levantamento de Recursos naturais, v.5.
PROJETO RADAMBRASIL. Folha SA.21 - Santarém. Belém: DNPM, 1976. v. 10. p. 310-414.
RICKLEFS, R. E. Ecology. 2. ed. New York: Chiron Press, 1979. p. 688-7.
RODRIGUES, W. A. Estudo de 2,6 ha de mata de terra firme da Serra do navio, Território do Amapá. Bol. Mus. Para. Emílio Goeldi, ser. Bot, Belém, v. 19, p. 1-44, 1963.
RUOLOLAINEN, K.; TUOMISTO, H. Beta-Diversity in Tropical Trees. Science, v. 297, p. 1439a, 2002.
SALOMÃO, R. P.; LISBOA, P. L. B. Análise ecológica da vegetação de uma floresta pluvial tropical de terra firme, Rondônia. Bol. Mus. Para. Emílio Goeldi, ser. Bot., Belém, v. 4, n. 2, p. 195-233, 1988.
SALOMÃO, R. P. Uso de parcelas permanentes para estudos de vegetação florestal I. Município de Marabá, Pará. Bol. Mus. Para. Emílio Goeldi, ser. Bot., Belém, v. 7, n. 2, p. 543-604, 1991.
SALOMÃO, R. P.; ROSA, N. A.; NEPSTAD, D. C.; BAKK, A. Estrutura diamétrica e breve caracterização ecológica-econômica de 108 espécies arbóreas da floresta amazônica brasileira. Interciência, v. 20, n. 1, p. 20-9, 1995.
SALOMÃO, R. P.; SILVA, M. M. F.; ROSA, N. A. Inventário ecológico em floresta pluvial tropical de terra firme, Serra Norte, Carajás, Pará. Bol. Mus. Para. Emílio Goeldi, ser. Bot., Belém, v. 4, n. 1, p. 1-46, 1988.
SCUDELLER, V. V.; MARTINS, F. R.; SHEPHERD, G. J. Distribution and abundance of arboreal species in the Atlantic Ombrophilous Dense Forest in Southeastern Brazil. Plant Ecology, Dordrecht, v. 152, n. 2, p. 185-99, 2001.
SHANLEY, P.; ROSA, N. A. Conhecimento em erosão: um inventário etnobotânico na fronteira de exploração da Amazônia oriental. Bol. Mus. Para. Emílio Goeldi, ser. Ciências Naturais, Belém, v. 1, n. 1, p. 147-71, 2005.
SHANNON, C. E.; WIENER, W. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1949. 117 p.
SILVA, M. F. F.; ROSA, N. A.; SALOMÃO, R. P. Estudos botânicos na área do Projeto Carajás. 3 - Aspectos florísticos da mata do aeroporto de Serra Norte - PA. Bol. Mus. Para. Emílio Goeldi, ser. Bot., Belém, v. 2, n. 2, p. 169-87, 1986.
SIMPSON, E. H. 1949. Measurement of diversity. Nature, v. 163, p. 688, 1949.
SIOLI, H. The Amazon: limnology and landscape ecology of a mighty tropical river and its basin. Dordrecht: W. Junk publishers, 1984. 763 p.
SöRENSEN. A method of establishing groups of equal amplitude in plant sociology based on similarityof species contents. Kong. Danske Vidensk. Selsk. Biol. Skr,, v 5, n. 4, p. 1-34, 1943.
STEEGE, H. Ter; PITMAN, N.; SABATIER, D.; CASTELLANOS, H.; DER HOUT P. V.; DALY D. C.; SILVEIRA, M.; PHILLIPS, O.; VASQUEZ. R.; ANDEL, T. V.; DUIVENVOORDEN, J.; OLIVEIRA, A. A.; RENSKE, E. K.; LILWAH, R.; THOMAS, R.; ESSEN, J. van; BAIDER, C.; MASS, P.; MORI, S.; TERBORGH, J.; VASGAR, P. N.; MOGOLLON, H.; WETZ, W. M. A Spatial Model of Tree -diversity and Tree Density for the Amazon. Biodiversity and Conservation, London, v. 12, p. 2255-77, 2002.
UHL, C., BUSCHBACHER, R.; SERRÃO, E. A. S. Abandoned pastures in eastern Amazonia. I. Patterns of plant succession. Journal of Ecology, v. 76, p. 663-81, 1988.
VELOSO, H. P.; RANGEL FILHO, A. L. R.; LIMA, J. C. Clasificação da vegetação brasileira, adaptada a um sistema universal. Rio de Janeiro: IBGE, Dirtetoria de Geociências, 1991. 123 p.
WHITMORE, T. C.; PRANCE, G. T. Biogeography and quaternary history in Tropical America. Oxford: Claredon Press, 1987. 212 p.
WRIGHT, J. F.; FURSE, M. T.; ARMITAGE, P. D.; MOSS, D. New procedures for identifying running-water sites subjects to environmental stress and for evaluating sites for conservation, based on the macroinvertebrate fauna. Archiv fur Hydrobiologie, Stuttgart, v. 127, n. 3, p. 319-326, 1993.
 Endereço para correspondência:
Endereço para correspondência:
Museu Paraense Emílio Goeldi
Editor do Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais
Av. Magalhães Barata, 376
São Braz - CEP 66040-170
Belém - PA - Brasil
Caixa Postal 399
Telefone/fax: 55-91- 3249 -1141
E-mail: boletim@museu-goeldi.br
Recebido: 17/05/05
Aprovado: 26/04/07
