










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.3 n.3 Belém dez. 2008
Escamas de tubarões (Pisces: Chondrichthyes) da Formação Pirabas (Eomioceno), Pará, Brasil
Shark scales (Pisces: Chondrichthyes) of Pirabas Formation (Eomiocen), Pará, Brazil
Samantha Florinda Cecim Carvalho de OliveiraI; Peter Mann de ToledoII; Sue Anne Regina Ferreira da CostaIII
IMuseu Paraense Emílio Goeldi. Belém, Pará, Brasil (samanthacecim@yahoo.com.br)
IIInstituto Nacional de Pesquisas Espaciais. São José dos Campos, São Paulo, Brasil (peter@dir.inpe.br)
IIIMuseu Paraense Emílio Goeldi. Belém, Pará, Brasil (sue.costa@gmail.com)
RESUMO
A falta de posicionamento estratigráfico dos fósseis de tubarões até então coletados na Formação Pirabas dificultou a utilização dos mesmos em interpretações paleoecológicas e distribuição temporal. Como forma de preencher esta lacuna em nosso conhecimento, foi utilizada a "screenwashing" em 14 amostras, estratigraficamente controladas, provenientes da Mina B17 (CIBRASA S/A), município de Capanema, que permitiu coletar e descrever, pela primeira vez, escamas de tubarões na unidade em questão. Ao todo, são 400 espécimes, divididos em sete categorias morfotípicas de acordo com o formato e topografias encontradas na coroa. Estas foram registradas em todos os níveis e podem ser atribuídas às ordens Orectolobiformes, Carcharhiniformes e Lamniformes, já descritas para a Formação Pirabas, e que corroboram o ambiente raso, marinho e tropical.
Palavras-chave: Formação Pirabas. Escamas. Tubarões.
ABSTRACT
The lack of stratigraphic localization of shark fossils collected in the Pirabas Formation has made it difficult to use them in paleoecological interpretation and distribution in time. As a way to fill this gap in our knowledge, the screenwashing technique was used in 14 samples, stratigraphically controlled and hailing from the B17 mine (CIBRASA S/A), in Capanema. This provided the first collection and description of shark scales. There are 400 specimens, which can be divided into seven morphotypes according to the format and topography found on the dental crown. These morphotypes were found in all levels and they belong to the orders Orectolobiformes, Carcarhiniformes and Lamniformes, that were already described from the Pirabas Formation and confirm the shallow, marine and tropical environment.
Keywords: Pirabas Formation. Scales. Sharks.
INTRODUÇÃO
Estudos que se propõem a enfocar aspectos paleobiogeográficos do Neógeno das regiões do Atlântico tropical e do mar do Caribe necessariamente têm que levar em consideração informações provenientes da paleofauna eomiocênica marinho-costeira da Formação Pirabas. Além da extensa lista de invertebrados fósseis conhecidos na literatura clássica sobre esta unidade geológica (Távora et al., 2004), mais recentemente, o volume de informações sobre a paleocomunidade de vertebrados também vem incrementando de forma significativa (Costa et al., 2004). Em particular, a ocorrência abundante de restos de tubarões em diferentes ambientes e estratos, e com grande diversidade taxonômica, fez destes peixes cartilaginosos uma importante fonte de informação para o entendimento de aspectos paleoecológicos/paleoambiental desta unidade estratigráfica (esta informação consta na Dissertação de Mestrado de S. A. F. Costa, do ano de 2005, intitulada "A comunidade de tubarões (Chondrichthyes: Selachii: Galea) da Formação Pirabas, Néogeno da Amazônia Oriental").
A caracterização taxonômica dos tubarões da Formação Pirabas e suas respectivas inferências paleoambientais foram efetivadas por Santos & Travassos (1960) e Santos & Salgado (1971), tendo sido revisadas recentemente por S. A. F. Costa e Reis (2005). Neste trabalho, utiliza-se a proposta de S. A. F. Costa, pois foi baseada em sinapomorfias mais eficazes, segundo a literatura mais moderna sobre o tema, e também apresenta o exame minucioso dos espécimes depositados no acervo de Paleontologia do Museu Paraense Emílio Goeldi. Assim, até o presente momento, foram reconhecidos: Carcharhinus spl, Carcharhinus sp2, Carcharhinus sp3, Carcharhinus priscus (Agassiz 1843), Galeoerdo sp., Sphyrna magna Cope 1867, Hemipristisserra Agassiz 1843, Carcharodon sp., Isurus sp., Nebrius obliquus (Leidy, 1877), Ginglymostoma serra (Leidy, 1877) e Carcharhininae indet.
Apesar da alta diversidade taxonômica e abundância dos tubarões, as técnicas de coleta utilizadas restringem-se à retirada de fragmentos macroscópicos da superfície rochosa, sem qualquer controle estratigráfico, que causa perda de valiosas informações sobre os padrões de distribuição temporal e ambiental ao longo das seções geológicas analisadas. A lacuna ocasionada pela falta dessas informações para a Formação Pirabas representa, hoje, o maior impedimento na utilização deste grupo de vertebrados em trabalhos de Estratigrafia (Rossetti & Góes, 2004).
Neste contexto, a utilização de material microscópico de tubarões torna-se uma fonte adicional e mais eficaz para a coleta de informações com enfoque no controle a padrões de abundância e ocorrência estratigráfica de vertebrados na Formação Pirabas. A prospecção de escamas vem a contribuir para o melhor entendimento da distribuição temporal e espacial destes vertebrados fósseis. Este trabalho complementa estudos anteriores (S. A. F. Costa) e detalha, pela primeira vez, o registro de escamas de tubarões da Formação Pirabas, além de contribuir para o refinamento das interpretações paleoambientais e paleoautoecológicas dos condrícteos.
Considerações Gerais sobre Escamas de Tubarões
As escamas de tubarões ou dentículos dérmicos têm por finalidade principal a proteção, sendo encontradas ao longo de toda extensão do corpo, incluindo a membrana nictitante, nadadeiras, cavidade oral, faringe e clásper (no caso dos machos). Esses dentículos dérmicos foram denominados de odontodes, a mesma identificação dada à estrutura originária ao esqueleto dermal (incluindo dentes) de todos os vertebrados, sendo que nos elasmobrânquios permaneceu pouco diferenciada ao longo do processo evolutivo (Sire & Huysseune, 2003). Porém, neste trabalho, decidiu-se pela utilização do termo escama, segundo nomenclatura de Reif (1985).
De forma geral, são consideradas placóides, pois não possuem uma lâmina de tecido ósseo nas camadas mais profundas, diferentemente do que se observa nas escamas dos peixes ósseos (ciclóides ou ctenóides). Assim como em relação ao seu crescimento, pois estas não crescem continuamente, são formadas durante um único processo (Reif & Richter, 2001). Estruturalmente, são divididas em: cavidade pulpar, dentina e, externamente, enamelóide (Springer & Gold, 1989). Este último é considerado como uma dentina modificada, formada pela ectoderme e mesênquima (Grady, 1970).
O padrão morfológico simples da escama de tubarão é constituído por uma base em forma de quadrado, sustentando a coroa (Reif, 1985). Os quatro cantos da base são chamados de processos. A coroa consiste de quatro "expansões" ('wings' sensu Reif, 1985), uma anterior, duas laterais e uma posterior. As "expansões", assim como os processos da base, seguem a orientação do corpo (Reif, 1985) (Figura 1A).
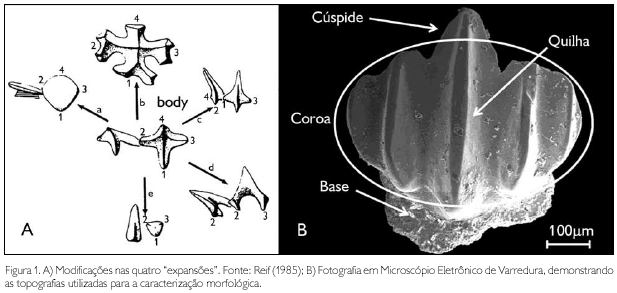
Os morfotipos foram definidos, neste trabalho, a partir das modificações na coroa segundo Reif (1985), ou seja, nas "expansões", e do posterior aparecimento de topografias, como cúspides e quilhas. As cúspides são proeminências na superfície da coroa que sofrem uma curvatura até se fundirem com a 4a expansão (Reif, 1985), enquanto as quilhas são dobras que podem estar em toda a superfície ou somente na borda da mesma (Thies, 1995) (Figura 1B).
A utilização de critérios morfológicos para definição de morfotipos de escamas de tubarões também foi realizada por Johns et al. (2005), porém os autores possuem uma abordagem bioestratigráfica do material, ou seja, não se detêm a detalhes criteriosos da superfície da coroa, já que todas são consideradas formas modernas e típicas do Cenozóico. Contudo, esses critérios definem as características morfuncionais das mesmas (Reif, 1985) e auxiliam nas interpretações paleoambientais.
Estas modificações na coroa são muito comuns em escamas, que, além de estarem diretamente ligadas à funcionalidade, também podem refletir estágios ontogenéticos (Reif, 1985). Por exemplo, se o exemplar apresentasse um baixo grau de mineralização, poder-se-ia inferir que o tubarão era relativamente jovem, ocorrendo um processo acumulativo de minerais com as diferentes fases de vida do indivíduo. No entanto, como esta característica ainda não pode ser corroborada por meio de análises em Microscópio Eletrônico de Varredura (Reif, 1985), optou-se utilizar para este trabalho apenas o aspecto morfológico como um parâmetro de discriminação entre as diferentes fases etárias.
MATERIAL E MÉTODOS
O trabalho de prospecção paleontológica foi desenvolvido em rochas pertencentes à Formação Pirabas, expostas mecanicamente na Mina B-17, localizada no município de Capanema, estado do Pará (Figura 2). Esta mina, aberta para exploração de calcário, pertence à empresa CIBRASA S/A, que detém o direito de lavra da pedreira (Távora et al., 2007).
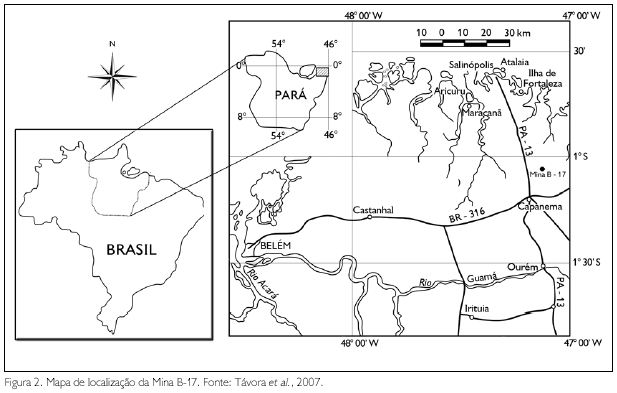
A Formação Pirabas ocorre predominantemente no litoral norte brasileiro, no estado do Pará, e está deposicionada na Plataforma Bragantina (Rossetti & Góes, 2004). A unidade é caracterizada pela presença de rochas carbonáticas que representam depósitos identificados como de plataforma externa (grainstone/ Pack Stone maciço, wackestone-packstone estratificado e argilito laminado), contendo também fácies litorâneas (shoreface/foreshore), além de elementos de plataforma restrita/laguna (marga, calcilutito, argilito cinza e verde oliva e biocalcarenito não estratificado) e de mangue (argilito negro, maciço ou laminado) (Rossetti, 2001).
Foram identificados 17 horizontes estratigráficos de uma única seção contendo, aproximadamente, 20 m de altura. Classificados litologicamente e identificados pela letra B seguida de um número. O perfil pode ser descrito resumidamente como possuindo calcários bem compactados (B0, B5 e B9), seguido por intercalações de calcarenitos de diferentes granulometrias (B1, B1A, B2, B2A, B3, B4, B7, B8, B10, B11 e B12); ainda nessa seqüência deposicional, ocorre uma camada argilosa (B6) e seu topo contém camadas de calcários intercaladas de argilito (B13). Neste trabalho, foram processados 14 níveis; as camadas B0 e B10 não foram amostradas devido à dificuldade de coletar sedimento suficiente em campo, e B14 é gradacional entre as Formações Pirabas 1. e Barreiras (Figura 3).
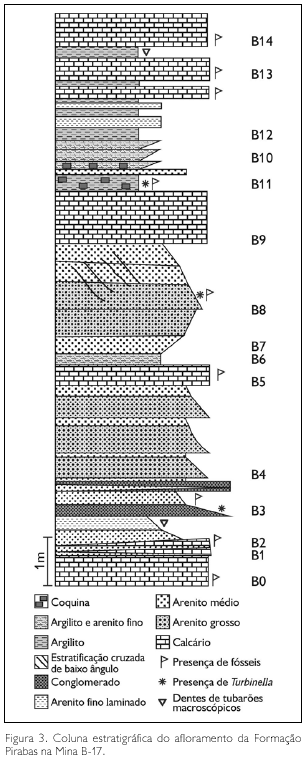
Foram coletados 4 kg de sedimentos de cada nível (B1, B1A, B2, B2A, B3, B4, B5, B6, B7, B8, B9, B11, B12 e B13); os materiais passaram por técnicas laboratoriais para 2. a recuperação de microfósseis de vertebrados, usando orientações adaptadas de Aldridge (1990), Johnson et al. (1994) e Thies (1995), onde ocorreu a desagregação da matriz por meio de imersão em ácido acético a 10%, por 3. 24 a 48 h, para o processo de 'screenwashing'.
Posteriormente, de cada nível foram retiradas subamostras de 230 g, retidas no intervalo de 60 'mesh', que foram triadas em microscópio estereoscópico. Depois, foram metalizadas com Au e fixadas em suportes para 4. Fotografia em Microscopia Eletrônica de Varredura de modelo LEO 1450 VP, do Laboratório Institucional de 5. Microscopia Eletrônica do Museu Paraense Emílio Goeldi.
RESULTADOS E DISCUSSÃO
A análise microscópica dos 14 níveis estratigráficos estudados rendeu 400 escamas de tubarões, cujas características morfológicas permitiram dividir o material 6. em sete diferentes morfotipos, descritos a seguir:
1. Escamas sem quilhas e sem cúspide: a coroa não possui ornamentações e o seu formato é arredondado, ocasionado pela fusão das quatro "expansões". Apêndice 1 (Figuras A-F);
2. Escamas sem quilhas e com cúspide: possuem a cúspide proeminente, apontada para parte posterior do animal e fundida com a quarta "expansão" da coroa. Apêndice 1 (Figuras G e H);
3. Escamas com quilhas em toda a superfície e sem cúspide: apresenta uma quilha principal, localizada medialmente na coroa, e laterais que podem ser de duas até seis em cada "expansão". Apêndice 2 (Figuras I-N);
4. Escamas com quilhas na extremidade anterior da coroa e sem cúspide. Apêndice 2 (Figuras O e P);
5. Escamas com quilhas em toda a superfície da coroa com cúspide: a cúspide está voltada para trás e fundida com a "expansão" posterior. Apresenta uma quilha principal mais desenvolvida, localizada medialmente, e outras laterais que podem ser de duas a três quilhas em cada "expansão". Apêndice 3 (Figuras Q-T);
6. Escamas com quilhas na extremidade anterior da coroa e presença de cúspide: a cúspide é direcionada para trás e fundida com a "expansão" posterior da coroa. Apêndice 3 (Figuras U-W);
7. Escama com quilhas e formação de ponta: a ponta está voltada para cima e para trás, porém não está fusionada com a quarta "expansão" posterior da coroa. Apêndice 3 (Figura X).
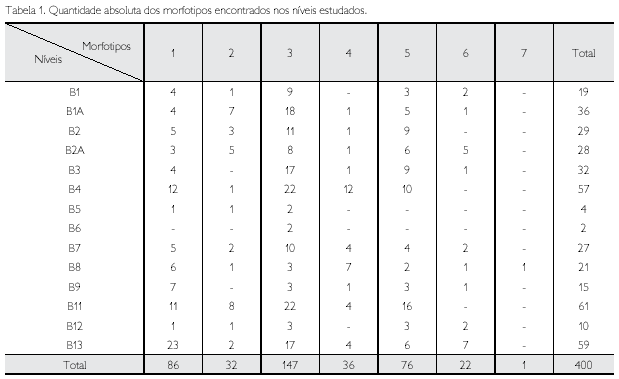
Entre as escamas encontradas na Formação Pirabas, o mais abundante é o morfotipo 3, com 147 exemplares distribuídos ao longo de todos os níveis. Contrariamente, o morfotipo 7 apresentou apenas uma ocorrência no nível B8. Como já esperado, mais de 90% da amostra deste estudo são compostas por escamas que podem pertencer a representantes de Carcharhinus (Reif, 1985), gênero mais diverso e abundante no registro fossilífero da unidade em questão (Costa et al., 2004), além de ser o melhor estudado em relação à morfologia de suas escamas (Raschi & Tabit, 1992) (Tabela 1).
Quase todos os morfotipos estudados podem ser atribuídos, de alguma forma, ao gênero Carcharhinus, recobrindo alguma parte do corpo em qualquer um dos seus estados ontogenéticos, com exceção do morfotipo 7. Este morfotipo ocorre apenas em embriões de Galeocerdo, na parte dorsal da cabeça (Reif, 1985).
Quanto à especificidade de ocorrência em embriões, quatro morfotipos podem pertencer a embriões de Carcharhinus, Galeocerdo e Sphyrna; são estes: 4, 5, 6 e 7. Estas escamas surgem gradualmente somente na segunda metade da embriogênese do animal. Porém, existem escamas típicas dessa fase que se diferenciarão ao nascer ou no crescer do indivíduo (Reif, 1985).
Diferentemente dos outros morfotipos que apresentam uma função mais de recobrimento corporal, viabilizando o melhoramento hidrodinámico (Raschi & Musick, 1986) do animal, o morfotipo 4 pode ser considerado como dentículo de revestimento, já que, em sua maioria, é encontrado nas fendas branquiais, cavidade oral e membrana nictitante dos gêneros Sphyrna e Carcharhinus (Reif, 1985).
Considerações Paleoambientais
As escamas de tubarões estão presentes em todo perfil estratigráfico (Tabela 1), sendo que no nível B11 são mais abundantes, com 61 exemplares, e, contrariamente, no nível B6, com apenas duas escamas. Por não serem diagnósticas de espécies, as interpretações paleoecológicas serão realizadas a partir da comparação anatômica das mesmas, utilizando o princípio da analogia, seguindo critérios de Reif (1985).
Uma das premissas para a utilização destes critérios é reconhecer que os tubarões apresentam variações em relação ao recobrimento corporal, ou seja, diferentes tipos de escamas ao longo dos estágios ontogenéticos (Reif, 1985). Isso se deve, principalmente, à otimização nos diferentes nichos ecológicos ao longo dos sucessivos estágios de vida (Carlson, 1998; Bethea et al., 2004). Entre os exemplares de escamas encontrados, a maior parte (66,25%) pode ser atribuída a juvenis e adultos. Esse resultado suporta a hipótese do ambiente marinho para a maioria dos níveis estudados, uma vez que os tubarões são comuns em ambientes transicionais, principalmente quando filhotes (Wetherbee & Cortés, 2004; Lowe et al., 1996).
É importante ressaltar que não ficou evidenciado, nos estudos, haver algum nível com predominância de escamas de embriões que pudesse indicar áreas como berçário. No entanto, analisando o comportamento de ocupação atual dos gêneros encontrados na Formação Pirabas (Figura 4), percebe-se que estes possuem hábitos semelhantes.
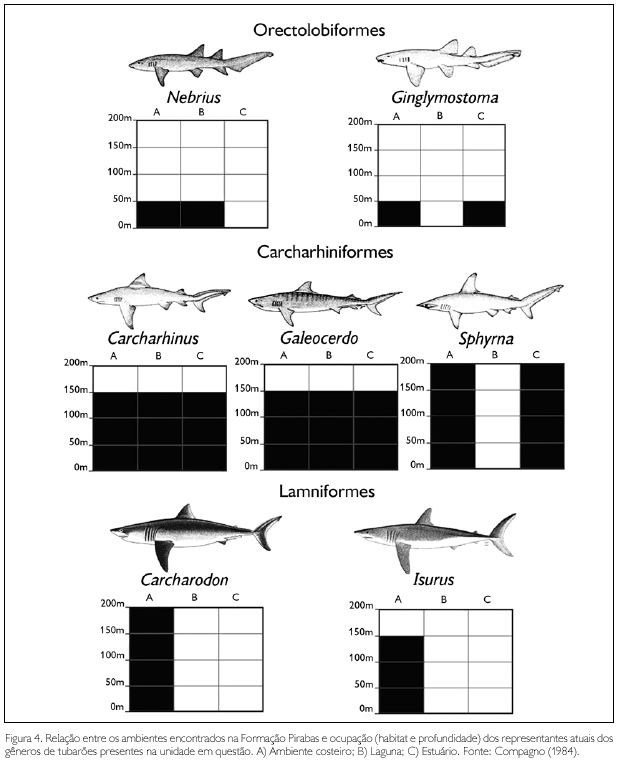
Como citado anteriormente, as escamas foram encontradas ao longo de todo perfil. E este, a partir da associação de suas fácies, apresenta todos os diferentes sub-ambientes já descritos para a unidade em questão, confirmando a hipótese de que os tubarões, quando juvenis e adultos, habitavam no Mioceno os ambientes marinhos, lagunar e estuarino (Compagno, 1984).
A paleocomunidade de tubarões da Formação Pirabas pode ser considerada como típica de ambiente tropical e costeiro, apresentando gêneros como Carcharhinus, não só o mais diverso em áreas costeiras, como também entre os tubarões. Por conta disto, suas espécies apresentam diferentes comportamentos em relação à profundidade, que, no caso, seria entre 18 m na costa, e algumas espécies alcançando até 500 m em mares abertos (Compagno, 1984). Como este trabalho limita-se ao estudo de gêneros, as escamas que podem ser atribuídas a este (morfotipos 1, 3, 4, 5 e 6) são encontradas em todos os níveis (Tabela 1). Deste modo, apesar de abundante, este grupo não contribui para a indicação batimétrica mais precisa (Figura 4).
Por outro lado, gêneros como Ginglymostoma e Nebrius não possuem grande variação, habitando áreas extremamente rasas, que se encontram entre 1 m a 30 m de profundidade (Compagno, 1984). As escamas que podem ser atribuídas a esses animais são os morfotipos 1, 2, 5 e 6, os quais ocorrem em todos os níveis, exceto em B6 (Tabela 1). Além do mais, estas ocorrências corroboram a variação batimétrica proposta de 0 a 50 m para a unidade, conforme já identificado entre a paleofauna de invertebrados (Ferreira, 1966).
Como já citado anteriormente, os gêneros Nebrius e Ginglymostoma, representantes da ordem Orectolobiformes (Shirai, 1996; Compagno, 1984), transitam em ambientes mais rasos e, segundo Reif (1985), estão dentro da categoria de tubarões demersais que vivem em substratos arenosos e lamosos. As escamas que representam esta categoria possuem a superfície da coroa lisa sem quilhas, conferindo-as como função de proteção maior do que uma relação de hidrodinámica. Conforme apresentado na Tabela 1, os morfotipos 1 e 2 se enquadram nesta categoria e ocorrem em quase todos os níveis prospectados.
A ordem mais diversa encontrada na Formação Pirabas é a Carchariniformes, cujos representantes são os gêneros Hemipristis, Galeocerdo, Sphyrna e Carcharhinus. Carcharhinus possui grande parte dos morfotipos descritos entre o material coletado, exceto morfotipos 2 e 7, que se apresentam em Sphyrna e Galeocerdo, respectivamente. Essa ordem é formada pelos tubarões considerados caçadores rápidos ou de velocidade moderada (Reif, 1985) e suas escamas estão adaptadas para reduzir a abrasão e proporcionar uma maior velocidade natatória ao animal (Raschi & Musick, 1986; Reif, 1978). A presença de quilhas contribui para uma maior estabilidade de movimento, uma vez que a presença de fendas longitudinais em "V" ao longo do corpo do tubarão diminui a abrasão por romper com as áreas de turbulência, aumentando em 10% a sua eficiência hidrodinâmica, se comparada com escamas de superfície lisa (Walsh & Weinstein, 1978). Com isso, a presença relativamente mais abundante dos morfotipos 3 e 5 coaduna-se com a predominância de representantes da família Carcharhinidae entre a paleocomunidade de tubarões em estudo.
A ordem Lamniforme representa os tubarões de tamanho avantajado (Compagno 1973), ou seja, os gêneros Carcharodon e Isurus. Estes animais são considerados predadores pelágicos rápidos (Reif, 1985) e de deslocamento por grandes distâncias (Dissertação de Mestrado de S. A. F. Costa; Aguilera & Rodrigues de Aguilera, 2004). Nas amostras, as escamas que provavelmente poderiam ser assinaladas como pertencentes a estes grupos são os morfotipos 2, 5 e 6.
O gênero Carcharodon estaria representado apenas pelo morfotipo 6. A razão de este gênero de importância na cadeia trófica da comunidade estar tão parcamente representado deve estar relacionada ao seu comportamento alimentar. Na estrutura trófica atual, o gênero Carcharodon ocupa uma das posições do topo da cadeia alimentar, tendo uma posição de consumidor terciário (Wetherbee & Cortés, 2004), além do fato de que eles habitavam áreas mais costeiras, apenas momentaneamente para alimentação (Aguilera & Rodrigues de Aguilera, 2004). A escassez relativa de escamas estaria diretamente ligada a uma menor freqüência de representantes dentro da paleocomunidade da Formação Pirabas, hipótese também corroborada pela menor incidência de elementos dentários do gênero (Costa et al., 2004).
CONCLUSÕES
A utilização de escamas de tubarões demonstrou ser uma ferramenta útil para estabelecer inferências sobre características paleoambientais e compará-las com informações publicadas sobre os aspectos geológicos e paleontológicos da Formação Pirabas. Algumas conclusões puderam ser alcançadas, por exemplo:
1) As escamas lisas, típicas de animais demersais, permitiram corroborar que a lâmina d'água do antigo mar de Pirabas, provavelmente, esteve limitada a 50 m de profundidade;
2) A presença de um número relativamente abundante de escamas com quilhas, feição esta característica de tubarões de velocidades moderadas e eficientes caçadores, encontrados principalmente em ambientes costeiros tropicais, como os representantes da família Carcharhinidae (ordem mais abundante e diversa), sugere que o ambiente deposicional era costeiro e de águas quentes;
3) A baixa representatividade de escamas atribuídas ao gênero Carcharodon, tubarão típico de ambiente profundo em relação aos Carcharhinus costeiros, pode refletir também questões de relativa baixa profundidade que predominava na região onde as incursões desta espécie no paleoambiente de Pirabas seriam esporádicas.
AGRADECIMENTOS
Os autores agradecem ao CNPq, pelo financiamento do projeto intitulado Paleoecologia e Bioestratigrafia da Formação Pirabas, nordeste do estado Pará, processo n° 480370/20044; ao Museu Paraense Emílio Goeldi, pela logística; à empresa CIBRASA S/A, por permitir o acesso à Mina B-17; à Dra. Dilce Rossetti, pela confecção do perfil; à Dra. Maria Inês Ramos, por auxiliar no início da pesquisa; e por fim, um agradecimento especial à Dra. Patrícia Charvet-Almeida, pesquisadora colaboradora do Museu Paraense Emílio Goeldi e SENAI-PR, pela disponibilização de bibliografias especializadas e revisão dos resultados aqui apresentados.
REFERÊNCIAS
AGUILERA, O. & D. RODRIGUES DE AGUILERA, 2004. Giant-toothed White Sharks and Wide-toothed Mako (Lamnidae) from the Venezuela Neogene: Their Role in the Caribbean, Shallow-water Fish Assemblage. Caribbean Journal of Science 40(3): 368-382.
ALDRIDGE, R. J., 1990. Extraction of Microfossils. In: D. E. F. BRIGGS & P. R. CROWTHER (Eds.): Paleobiology - a Synthesis: 502-504. Blackwell Scientific Publications, Oxford.
BETHEA, D. M., J. A. BUCKEL & J. K. CARLSON, 2004. Foraging ecology of the early life stages of four sympatric shark species. Marine Ecology Progress Series 268: 245-264.
CARLSON, J. K., 1998. Occurrence of neonate and juvenile sandbar sharks, Carcharhinusplumbeus, in the northeastern Gulf of Mexico. Fishery Bulletin 97(2): 387-391.
COMPAGNO, L. J. V., 1984. Sharks ofthe World. An annotaded and illustraded catalogue of sharks species know to date. FAO species catalogue 4(1, 2): 1-655.
COSTA, S. A. F., P. M. TOLEDO & H. M. MORAES-SANTOS, 2004. Paleovertebrados. In: D. F. ROSSETTI & A M. GÓES (Eds.): Néogeno da Amazônia Oriental: 135-166. Museu Paraense Emílio Goeldi, Belém.
FERREIRA, C. S., 1966. Características lito-paleontológicas da Formação Pirabas, Estado do Pará. VI Conferência das Guianas 6: 101-111.
GRADY, J. E., 1970. Tooth development in sharks. Archives Oral Biology 15: 613-619.
JOHNS, M. J., C. R. BARNES & Y. R. NARAYAN, 2005. Cenozoic and Cretaceous Ichthyoliths from the Tofino Basin and Western Vancouver Island, British Columbia, Canadá. Palaeontologia Electronica 8(2): 1-202.
JOHNSON, G. D., P. A. MURRY & J. E. STORER, 1994. Recovery of Vertebrate Microfossil. Proceedings of the South Dakota Academy of Science 73: 211-230.
LOWE, C. G., B. M. WETHERBEE, G. L. CROW & A. L. TESTER, 1996. Ontogenetic dietary shifts and feeding behavior of the tiger shark, Galeocerdocuvier, in Hawaiian waters. Environmental Biology of Fishes 47: 203-211.
RASCHI, W. G. & J. A. MUSICK, 1986. Hydrodynamic Aspects of Sharks Scales: 1-116. National Aeronautics and Space Administration, Washington.
RASCHI, W. & C. TABIT, 1992. Functional Aspects of Placoid Scales: a review and update. Australian Journal of Marine & Freshwater Research 43: 123-147.
REIF, W. E., 1985. Squamation and Ecology of Sharks. Courier Forschungsinstitut Senckenberg 78: 1-255.
REIF, W. E., 1978. Protective and hydrodynamic function of the dermal skeleton of elasmobranches. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 157: 131-141.
REIF, W. E. & M. RICHTER, 2001. Revisiting the lepidomorial and the odontode regulation theories of dermo-skeletal morphogenesis. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 219: 285-304.
REIS, M. A. F., 2005. Chondrichthyan Fauna from the Pirabas Formation, Miocene of Northern Brazil, with Comments on Paleobiogeography. Anuário do Instituto de Geociências – UFRJ 28(2): 31-58.
ROSSETTI, D. F., 2001. Late Cenozoic sedimentary evolution in northeastern Pará, Brazil, within the context of sea level changes. Journal of South American Earth Sciences 14: 77-89.
ROSSETTI, D. F. & A. M. GÓES, 2004. Geologia. In: D. F. ROSSETTI & A. M. GÓES (Eds.): Néogeno da Amazônia Oriental: 13-52. Museu Paraense Emílio Goeldi, Belém.
SANTOS, R. S. & M. S. SALGADO, 1971. Contribuição à Paleontologia do Estado do Pará - Novos restos de peixes da Formação Pirabas. Boletim do Museu Paraense Emílio Goeldi, Nova Série, Geologia 16: 1-13.
SANTOS, R. S. & S. TRAVASSOS, 1960. Contribuição à Paleontologia do Estado do Pará. Peixes fósseis da Formação Pirabas. Monografia da divisão de Geologia e Mineralogia, Departamento Nacional da Produção Mineral 16: 1-35.
SHIRAI, S., 1996. Phylogenetic interrelationships of neoselachians (Chondrichthyes: Euselachii). In: M. L. J. STIASSNY, L. R. PARENTI & G. D. JOHNSON (Eds.): Interrelationships of Fishes: 9-32. Academic Press, New York.
SIRE, J. Y. & A. HUYSSEUNE, 2003. Formation ofskeletal and dental tissues in fish: A comparative and evolutionary approach. Biological Reviews of the Cambridge Philosophical Society 78: 219-249.
SPRINGER, V. G. & J. P. GOLD, 1989. Sharks in question. The Smithsonian answer book: 1-187. Smithsonian Institution Press, Washington.
TÁVORA, V. A., E. S. F. SILVEIRA, J. M. MILHOMEM NETO, 2007. Mina B-17, Capanema, PA - Expressivo Registro de uma Paleolaguna do Cenozóico Brasileiro. In: M. WINGE, C. SCHOBBENHAUS, M. BERBERT-BORN, E. T. QUEIROZ, D. A. CAMPOS, C. R. G. SOUZA & A. C. S. FERNANDES (Eds.): Sítios Geológicos e Paleontológicos do Brasil. Disponível em: <http://www.unb.br/ig/sigep/sitio121/sitio121.pdf>. Acesso em: 17 novembro 2007.
TÁVORA, V. A., E. F. F. IMBELONI, A. S. M. CACELA & N. B. BAIA, 2004. Paleoinvertebrados. In: D. F. ROSSETTI & A. M. GÓES (Eds.): O Néogeno da Amazônia Oriental: 111-131. Museu Paraense Emílio Goeldi, Belém.
THIES, D., 1995. Placoid Scales (Chondrichthyes: Elasmobranchii) from the late Jurassic (Kimmeridgian) of northern Germany. Journal of Vertebrate Paleontology 15(3): 463-481.
WALSH, M. J. & L. M. WEINSTEIN, 1978. Drag and heat transfer on surfaces with small longitudinal fins. In: XI Fluid and Plasma Dynamics Conference: 1-12. American Institute of Aeronautics and Astronautics, Seattle.
WETHERBEE, B. M. & E. CORTÉS, 2004. Food Consumption and Feeding Habits. In: J. C. CARRIER, J. A. MUSICK & M. R. HEITHAUS (Eds.): Biology of Sharks and their Relatives: 223-242. Chemical Rubber Company Press, Boca Raton.
 Endereço para correspondência:
Endereço para correspondência:
Museu Paraense Emílio Goeldi
Editor do Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais
Av. Magalhães Barata, 376
São Braz – CEP 66040-170
Belém - PA - Brasil
Caixa Postal 399
Telefone: 55-91-3182-3246
Fax: 55-91-3249-6373
E-mail: boletim.naturais@museu-goeldi.br
Recebido: 16/01/2008
Aprovado: 26/12/2008
