










Serviços Personalizados
Journal

Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.4 n.3 Belém dez. 2009
Shark teeth from Pirabas Formation (Lower Miocene), northeastern Amazonia, Brazil
Dentes de tubarões da Formação Pirabas (Mioceno Inferior), Amazônia Oriental, Brasil
Sue Anne Regina Ferreira CostaI; Martha RichterII; Peter Mann de ToledoIII; Heloisa Maria Moraes-SantosIV
IMuseu Paraense Emílio Goeldi. Belém, Pará, Brasil (sue.costa@gmail.com)
IIThe Natural History Museum. Department of Palaeontology. London, United Kingdom (M.Richter@nhm.ac.uk)
IIIInstituto Nacional de Pesquisas Espaciais. São José dos Campos, São Paulo, Brasil (peter@dir.inpe.br)
IVMuseu Paraense Emílio Goeldi. Belém, Pará, Brasil (hmoraes@museu-goeldi.br)
ABSTRACT
The shark fauna (Chondrichthyes: Elasmobranchii) of Pirabas Formation is one of the most representative and diversified of the Neogene of South America. This fauna is associated with the early Miocene transgression resulting from the global sea level rise. In this paper, the composition of the Pirabas shark fauna is re-evaluated based on new fossil findings. The occurrence of the following taxa is confirmed: Carcharhinus sp.1, Carcharhinus sp.2, Carcharhinus cf. macloti, Carcharhinus priscus, Galeocerdo sp., Sphyrna cf. media, Hemipristis serra, Carcharodon subauriculatus, Isurus sp., Ginglymostoma sp., Nebrius obliquus.
Keywords: Pirabas Formation. Chondrichthyes. Neogene. Amazonia.
RESUMO
A fauna de tubarões (Chondrichthyes: Elasmobranchii) da Formação Pirabas é uma das mais representativas e diversificadas do Neógeno da América do Sul. Ela está associada à transgressão ocorrida no início do Mioceno, em consequência do aumento global do nível do mar. Neste trabalho, a composição da fauna de tubarões previamente identificada para a Formação Pirabas é reavaliada com base em novos achados fósseis. É confirmada a ocorrência dos seguintes táxons: Carcharhinus sp.1, Carcharhinus sp.2, Carcharhinus cf. macloti, Carcharhinus priscus, Galeocerdo sp., Sphyrna cf. media, Hemipristis serra, Carcharodon subauriculatus, Isurus sp., Ginglymostoma sp., Nebrius obliquus.
Palavras-chave: Formação Pirabas. Chondrichthyes. Neógeno. Amazônia.
INTRODUCTION
The lower Miocene Pirabas Formation occurs within coastal sedimentary basins between the states of Piauí and Pará, Northern Brazil. The best outcrops are found in the northwestern coast of the State of Pará, especially in the Salgado Zone region. Deposits of this formation include mostly carbonates intercalated with black and greenish shales and, secondarily, yellow sandstones (Rossetti & Góes, 2004). These rocks were formed under a variety of palaeoenvironmental conditions, including outer and inner shelf as well as tidal channel, lagoon, tidal and mangrove associated with barrier island systems. These depositional settings allowed the establishment of a diverse and abundant palaeofauna (Góes et al., 1990).
The fossiliferous assemblage of the Pirabas Formation has been described since the end of the 19th century and most of the studies have focused on invertebrates (e.g. Maury, 1925; Ferreira & Cunha, 1957; Távora, 1998), including microfossils (Távora, 1998; Ramos et al., 2004). Due to the fact that they are less commonly found, the vertebrates have received less attention. They are represented by sirenians (Toledo & Domning, 1989), fishes (Santos & Travassos, 1960; Costa et al., 2004) and, less frequently, crocodilians, chelonians and birds (Ackermann, 1964).
Among the vertebrates, fishes, which comprise osteichthyans and chondrichthyans, represent the most abundant and diverse group, showing the largest temporal and palaeogeographic distribution (Costa et al., 2004). This raises their potential as palaeoenvironmental and biostratigraphic proxies. The Pirabas fish fauna was firstly studied by Santos & Travassos (1960), who described nine shark species: Carcharhinus ackermannii, C. egertoni, Hemipristis serra, Scoliodon taxandriae, Galeocerdo paulinoii, Sphyrna magna, S. prisca, Ginglymostoma serra and G. obliquum. Later, large lamnid forms as Carcharodon megalodon and Isurus nova, and the genus Hipoprion were included (Santos & Salgado, 1971). More recently, Reis (2005) recognized a total of 13 taxa for the Pirabas Formation, ten of them representing extant taxa.
The chondrichthyan record from the Pirabas Formation is commonly restricted to isolated teeth and scales. In particular, the identification of elasmobranch fauna has been based on relatively small number of teeth (Santos & Travassos, 1960; Santos & Salgado, 1971; Reis, 2005). Based on knowledge about intraspecific variation of extant sharks, it becomes evident that a significant number of teeth is necessary for a precise taxonomic determination. The study of the dental morphological patterns in modern species has revealed a high degree of heterodonty in both jaws. Consequently, determinations based on isolated teeth can lead to misidentifications (Purdy et al., 2001).
In the last few years, systematic fieldwork in the Pirabas Formation led by researchers at the Museu Paraense Emílio Goeldi (MPEG) resulted in the addition of 289 new shark teeth to the MPEG paleontological collection. This represents a significant increase in the number of specimens when compared to materials described by Santos & Travassos (1960), Santos & Salgado (1971) and Reis (2005). This work aims to re-evaluate the shark taxa present in the Pirabas Formation, using shark teeth fossils deposited in the MPEG and from the collection of the Departamento Nacional de Produção Mineral (DNPM), in the state of Rio de Janeiro.
RESULTS AND DISCUSSION
Class Chondrichthyes
Subclass Elasmobranchii
Superorder Galea
Order Carcharhiniformes
Family Carcharhinidae
Subfamily Carcharhininae
Carcharhinus Blainville, 1816
Carcharhinus sp.1 (Santos & Travassos, 1960)
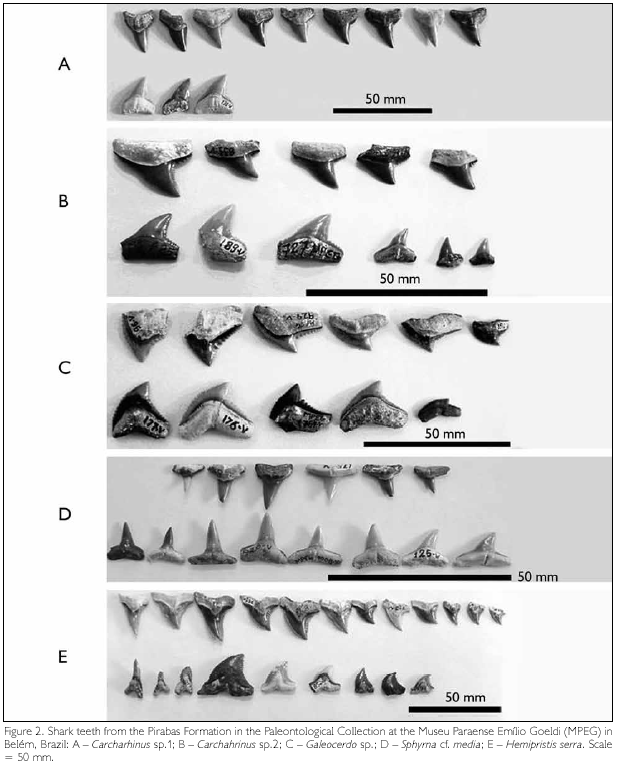
(Figure 1A, Figure 2A)

Carcharhinus ackermannii Santos & Travassos, 1960, p. 4, pl. 1, fig. 8-10.
Material: MPEG - 94-V, 109-V, 113-114-V, 124-V, 130-131-V, 155-157-V, 160-V, 178-V, 180-V, 183-V, 189-V, 190-V, 192-194-V, 274-277-V, 726-V, 729-730-V, 773-V, 786-V, 788-792-V, 797-V, 815-826-V, 828-829-V, 859-860-V, 907-V, 920-921-V, 937-V, 951-952-V, 954-V, 977-980-V, 988-V, 1032-V, 1033-V, 1095-V, 1097-V, 1106-1109-V, 1131-1133-V e DGM 651-P (holotype).
Dental diagnose: Crown triangular, unicuspidated, completely serrated with strong basal serration and an either deep or shallow basal notch. The teeth are straight in labial view and slightly convex in lingual view. Anterior teeth show straight and larger crowns, compared to posterior teeth. The holoaulacorhize root is flat with a central foramen.
Discussion: Diagnostic characters of Carcharhinus are only found on anterior upper teeth (Naylor & Marcus, 1994). The diagnosis provided for C. ackermannii (Santos & Travassos, 1960), contains characters currently only utilized to define the genus, except for the presence of small notches on the crown margin. The examination of 73 teeth has shown that such notches can not be considered diagnostic features. They are not uniform and vary in depth (Figure 1A, Figure 2A), thus they do not sustain the assignment to a species.
Carcharhinus sp.2
(Figure 1B, Figure 2B)
Material: MPEG - 100-V, 184-V, 198-V, 772-V, 785-V, 827-V, 830-835-V, 906-V, 919-V, 1064-V, 1154-1156-V.
Dental diagnose: Crown triangular, unicuspidated, completely serrated, with strong serration at the base, and lack of any notches. In the anterior tooth rows, the crowns are straighter than those in posterior rows. The teeth are straight in labial view and slightly convex linguad. The root is flat, holoaulacorhize and with a central foramen.
Discussion: C. egertoni is one of the Carcharhinus species most frequently cited in the literature. However, its distribution in the fossil record is highly contentious due to the difficulty of identifying isolated teeth (Cappetta, 1987). Reassessment of material from the Neogene of North Carolina, USA, demonstrated that the material previously referred to C. egertoni in fact comprises three species, namely C. brachyurus, C. falciformis e C. leucas (Purdy et al., 2001).
Previous identification of C. egertoni in the Pirabas Formation was based on two isolated teeth and their description by Santos & Travassos (1960) included only generic characters. Considering that the re-examined teeth occupied a lateral position on the jaw as indicated by their posteriorly inclined crowns, they cannot be ascribed to a particular species. As noted by Naylor & Marcus (1994), only anterior teeth bear characters diagnostic of Carcharhinus species.
Carcharhinus cf. macloti (Müller & Henle, 1839)
(Figure 1C)
Material: MPEG - 923-V, 984-V, 1031-V and DGM 969-P.
Dental diagnose: Small tooth (3-7 mm in height), crown triangular, unicuspidated, with the central portion of the cusp smooth and strong basal serrations. The teeth are straight in labial view and slightly convex lingually. Holoaulacorhize root with a central foramen.
Discussion: This specimen resembles the C. macloti documented in the Miocene (Pungo River Formation) and lower Pliocene (Yorktown Formation) deposits of USA (Purdy et al., 2001). Regardless of the scarcity of material (i.e., only two teeth), the characteristic strong serrations at the base of the crown and the smoothness of the central, higher portion of the crown suggest the presence of this species in the Pirabas Formation.
Carcharhinus priscus (Agassiz, 1843)
(Figure 1D, Figure 2C)
Material: MPEG - 93-V, 101-V, 102-V, 104-V, 110-V, 112-V, 117-V, 119-V, 127-V, 132-V, 159-V, 162-163-V, 164-165-V, 167-168-V, 170-V, 173-V, 186-V, 771-V, 773-774-V, 795-796-V, 848-851-V, 914-V, 933-936-V, 950-V, 975-V, 1013-V, 1034-1036-V, 1081-1082-V, 1140-1142-V, 1153-V and DGM 656-P.
Dental diagnose: Crown triangular and fine, unicuspidated, with uniform serrations along the entire border. The crowns in the anterior rows are straight and larger than those on posterior rows. Teeth straight in labial view and slightly convex lingually. Holoaulacorhize root with a central foramen.
Discussion: Carcharhinus priscus, together with C. egertoni, are some of the most cited species in the shark tooth literature. Carcharhinus priscus can easily be mistaken as representatives of the genus Sphyrna (Cappetta, 1987), as seems to have happen initially with materials from Pirabas Formation described by Santos & Travassos (1960). Although only two teeth were utilized in their descriptions, those authors had already pointed out differences between the teeth from the Pirabas Formation and those figured by Leriche (1927). Studying Miocene sharks from Ecuador, Longbottom (1979) referred the material from Pirabas Formation to Carcharhinus priscus.
Galeocerdo Müller & Henle, 1837
Galeocerdo sp.
(Figure 1E, Figure 2D)
Galeocerdo paulinoii Santos & Travassos, 1960, p. 8, pl. 1, fig. 13-14.
Material: MPEG - 95-96-V, 121-V, 146-V, 149-150-V, 176- 177-V, 199-V, 784-V, 857-V, 861-V, 929-V, 930-V, 931-V, 943-V, 1101-1103-V and DGM 106-P (holotype), DGM 653-P.
Dental diagnose: Crown triagular, unicuspidated, curved posteriorly. Mesial border convex, elongated and slightly sigmoidal. Crown strongly serrated with secondary serrations only in the median and basal parts of the crown. Holoaulacorhize root with a central foramen.
Discussion: Based on four complete and some fragmentary teeth, Santos & Travassos (1960) erected an endemic species for the Pirabas Formation. Nevertheless, the diagnosis presented by those authors contains only generic characters and even with the new collected material, it was not possible to confirm the presence of G. paulinoii in this formation.
On the other hand, features as tooth morphology, size of the principal cusp and type of compose serrations (Cigala-Fulgosi & Mori, 1979) show that the tooth from the Pirabas Formation could be ascribed to G. cuvier. However, these characters can also be found in the Miocene species G. aduncus (Purdy et al., 2001). As distinction between these two species can not be safely determined, this material is only identified at generic level herein.
Sphyrna Rafinesque, 1810
Sphyrna cf. media Springer, 1940
(Figure 1F)
Material: MPEG - 111-V, 118-V, 120-V, 122-123-V, 125- 126-V, 169-V, 171-V, 174-175-V, 181-V, 187-V, 278-V, 775-778-V, 821-V, 837-844-V, 846-847-V, 914-915-V, 925-V, 927-928-V, 976-V, 987-V, 1008-1009-V, 1059-V, 1063-V, 1066-V, 1068-V, 1069-V, 1077-V, 1104-1105-V, 1143-1147-V, 1149-1152-V.
Dental diagnose: Crown triangular, unicuspidated, slightly curved towards the labial side and without serrations. Teeth on anterior rows with straight crowns and larger than posterior ones. Flat, holoaulacorhize root with a central foramen.
Discussion: The diagnostic criteria of the genus Sphyrna can, in some cases, be misleadingly attributed to either S. media or other members of the subfamily Carcharhininae, mostly Rhizoprionodon. However, among extant representatives of these two genera, the crown is small, less then 4 mm high in Rhizoprionodon, while in Sphyrna the same measure is 1 to 2 cm (Cappetta, 1987). The crown height is, thus one of the criteria for the identification of the material from Pirabas Formation. In addition, the crown with either totally smooth borders or soft crenulations on the basal expansions are features typical of Sphyrna (Purdy et al., 2001). These characteristics led to relate the studied material in the Pirabas Formation to S. cf. media.
Carcharhininae indet.
Material: MPEG - 182-V, 188-V, 197-V, 787-V, 811-V, 836-V, 845-V, 858-V, 908-909-V, 924-V, 926-V, 955-V, 1014- V, 1037-V, 1062-V, 1078-V, 1157-V and DGM 654-655-P.
Dental diagnose: Crown triangular, unicuspidated, smooth and straight, showing only soft serrations at the base. Teeth in anterior rows show straight crowns that are larger than teeth on posterior rows. Root holoaulacorhize with a central foramen.
Discussion: Teeth belonging to this subfamily were ascribed to Scoliodon (Santos & Travassos, 1960). A review of the genus led to assign the Scoliodon species to three genera, including Rhizoprionodon, Loxodon and Scoliodon (Springer, 1964). This author was pessimistic about the possibility to distinguish between these genera based only on isolated fossil teeth. The close similarity between the teeth of Rhizoprionodon and Scoliodon and the lack of knowledge about the dental variations of living representatives of these genera hinder the determination of palaeontological material.
Compagno (1984) adopted the same diagnoses for Scoliodon and Rhizoprionodon, stressing the absence of serrations. However, a detailed study of complete dentitions of extant Rhizoprionodon shows that they bear either completely smooth teeth or teeth whose crown bases show various degrees of serrations (according to U. L. Gomes, in his Master Dissertation, entitled 'A dentição como um subsídio taxonómico dos Pleurotremata (Pisces, Chondrichthyes, Elasmobranchii)", defended in 1988). This finding precludes the secure identification at species level of isolated teeth.
Subfamily Hemigaleinae
Hemipristis Agassiz, 1843
Hemipristis serra Agassiz, 1843
(Figure 1G, H; Figure 2E)
Material: MPEG - 106-108-V, 145-V, 147-148-V, 153-V, 179-V, 273-V, 279-280-V, 724-725-V, 738-V, 779-783-V, 785-V, 793-794-V, 852-856-V, 905-V, 916-918-V, 922-V, 938-941-V, 1012-V, 1028-1030-V, 1055-V, 1099-V, 1100-V, 1134-1139-V.
Dental diagnose: Upper teeth with labio-lingually compressed, triangular crowns, strongly serrated. Lower teeth with hook-like, sharp crowns with narrow roots.
Discussion: Hemipristis serra is one of the commonest shark species in Miocene deposits and its strong dignatic heterodonty has led to erroneous identification of fossil materials (Cappetta, 1987). The dentition of the extant Hemipristis is well known, which facilitates precise identification of isolated teeth (Longbottom, 1979; Cappetta, 1987; Iturralde-Vinent et al., 1996; Purdy et al., 2001). Although the teeth of H. serra are similar to those of the extant species H. elongatus, studies have shown that the extinct species has smaller teeth and that sharks of this genus progressively increased the size of their teeth through geological time (Chandler et al., 2006). This information was taken into consideration for the identification of the material from Pirabas Formation. Additionally, based on recent and fossil material (S. A. F. Costa, personal observation), Hemipristis teeth from this unit are too large to represent juvenile specimens and too small to match the size of teeth of extant adult sharks of this genus. These characteristics further support the attribution of this material to H. serra.
Family Alopiidae
Subfamily Lamninaea
Carcharodon Smith in Müller & Henle, 1838
Carcharodon subauriculatus Agassiz, 1839
(Figure 1I)
Material: MEPG - 97-99-V, 154-V, 224-V, 723-V, 907-V, 1004-V and DGM 967-P.
Dental diagnose: Large teeth up to 840 mm in height, crown triangular, cusplets separated from the principal cusp by a slight notch. Holoaulacorhize root with rounded lateral lobes.
Discussion: The teeth from the Pirabas Formation belonging to this species were initially misidentified as C. megalodon (Santos & Salgado, 1971, p. 2). This was probably due to the fact that teeth of juvenile C. megalodon are reduced in size and also present lateral cusplets as C. subauriculatus. However, only C. subauriculatus possesses the distinctive notch separating the principal cusp from the lateral ones (Purdy et al., 2001). Additionally, C. megalodon remains are not known from rocks older than the middle Miocene (Applegate & Espinosa-Arrubarrena, 1996) and the Pirabas deposits belong to the early Miocene.
Isurus Cuvier, 1816
Isurus sp.
Material: MPEG - 932-V, 910-V e DGM 968-P.
Dental diagnose: Teeth with narrow crowns, higher than longer and without serrations or lateral cusplets. Teeth sigmoidal in lateral view. Bulky holoaulacorhize root with prominent lingual side.
Discussion: Identification of this genus in the Pirabas Formation was based on the presence of symphisial teeth (Santos & Salgado, 1971). However, these authors concluded that it was not possible to ascribe the teeth to a particular species within this genus. The new material is also fragmentary and insufficient to determine the Isurus species they belong to.
Family Rhincodontidae
Ginglymostoma Müller & Henle, 1837
Ginglymostoma sp.
(Figure 1J)
Material: MPEG - 813-V, 1073-V e DGM 657-P.
Dental diagnose: Teeth longer than higher, symmetrical with the lower part of the crown overhanging the root and forming a short protuberance. There is one principal central cusp with both sides presenting up to seven lateral cusplets. The root is hemiaulacorhize and flat.
Discussion: Teeth of Ginglymostoma, resemble those of Nebrius, which belongs to the same family. These are differentiated because in the latter the shorter cusps are linked together at their bases, forming a high platform. Additionally, Ginglymostoma bears seven lateral cusplets either side of the principal cusp, whilst in Nebrius there are eight cusplets either side (Cappetta, 1987).
The species G. serra was described in the Pirabas Formation on the basis of one single tooth (Santos & Travassos, 1960, p. 12) that does not show any characters that would allow a specific identification. The additional available specimens also do not present diagnostic features other than generic ones.
Nebrius Ruppell, 1837
Nebrius obliquus (Leriche, 1942)
(Figure 1K)
Material: MPEG - 814-V.
Dental diagnose: Teeth longer than higher, symmetrical with the base of the crown overhanging the root and forming a short protuberance. Median cusp not distinct from the lateral cusplets, which sum eight on either side. The root is hemiaulacorhize and flat.
Discussion: This species was identified as Ginglymostoma obliquum (Leidy, 1877) on the basis of two lateral teeth displaying middle cusp bending towards one of the crown margins (Santos & Travassos, 1960, p.13). However, this is also one of the marked characters of Nebrius (Bourdon & Heim, 2004). The only specimen of Nebrius in the paleontological collection of the MPEG is a fragmentary tooth, but it still shows diagnostic features as symmetrical crown and eight cusplets in either side of the middle cusp (see Purdy et al., 2001).
CONCLUSIONS
The following taxa were confirmed in the Pirabas Formation after a revision of previously described materials, as well as 289 new shark teeth recovered during recent fieldwork in the State of Pará, Northern Brazil:
Order Carcharhiniformes
Family Carcharhinidae
Subfamily Carcharhininae
Carcharhinus sp.1
Carchahrinus sp.2
Carcharhinus cf. macloti
Carcharhinus priscus
Galeocerdo sp.
Sphyrna cf. media
Carcharhininaea indet.
Subfamily Hemigaleinaea
Hemipristis serra
Order Lamniformes
Family Alopiidae
Subfamily Lamninaea
Carcharodon subauriculatus
Isurus sp.
Order Orectolobiformes
Family Rinchodotidae
Ginglymostoma sp.
Nebrius obliquus
ACKNOWLEDGMENTS
We thank an anonymous reviewer for the suggestions that improved the final version of this paper. We are also grateful to Sidclay Dias and Denys José Xavier Ferreira for capturing and processing the images, Marise Sardenberg for the assistance at the DNPM collection, and CNPq for supporting this research.
REFERENCES
ACKERMANN, F. L., 1964. Geologia e fisiografia da Região Bragantina (Estado do Pará). Cadernos da Amazônia (INPA) 2: 1-90.
APPLEGATE, S. & L. ESPINOSA-ARRUBARRENA, 1996. The fossil history of Carcharodon and its possible ancestor, Cretolamna: A study in tooth identification. In: A. P. KLIMLEY & D. G. AINLEY (Eds.): The Biology of the white shark, Carcharodon carcharhias: 19-36. Academic Press San Diego, Califórnia.
BOURDON, J. & B. HEIM, 2004. Extant Dentition. Available in: <http://www.elasmo.com>. Accessed on: 20 March 2004.
CAPPETTA, H., 1987. Condrichthyes II. Mesozoic and Cenozoic Elasmobranchii. Handbook of Paleoichthyology: 3B: 1-196. Gustav Fischer Verlag, Stuttgart.
CHANDLER, R., K. E. CHISWELL & G. D. FAULKNER, 2006. Quantifying a possible Miocene phyletic change in Hemipristis (Chondrichthyes) Teeth. Palaeontologia Electronica 9(1): 1-14.
CIGALA-FULGOSI, F. & D. MORI, 1979. Osservazioni tassonomiche sul genere Galeocerdo (Selachii, Carcharhinidae) (Peron e Lesueur) nel Pliocene del Mediterraneo. Bolletino della Società Paleontologica Italiana 18(1): 117-132.
COMPAGNO, L. J. V., 1984. Sharks of the world. An annotaded and illustrated catalogue of shark species know to date. FAO species catalogue 4(1/2): 1-655.
COSTA, S. A. F., P. M. TOLEDO & H. M. MORAES-SANTOS, 2004. Paleovertebrados. In: D. F. ROSSETTI & A. M. GÓES (Eds.): Neógeno da Amazônia Oriental: 135-166. Museu Paraense Emílio Goeldi, Belém.
FERREIRA, C. S. & O. R. CUNHA, 1957. Contribuição à Paleontologia do estado do Pará. Notas sobre a Formação Pirabas, com descrição de novos invertebrados fósseis (Mollusca– Gastropoda). Boletim do Museu Paraense Emílio Goeldi, Nova Série, Geologia 2: 1-60.
GÓES, A. M., D. F. ROSSETTI, A. C. R. NOGUEIRA & P. M. TOLEDO, 1990. Modelo deposicional preliminar para a Formação Pirabas, nordeste do Estado do Pará. Boletim do Museu Paraense Emílio Goeldi, série Ciências da Terra 2(1): 3-15.
ITURRALDE-VINENT, M., G. HUBBELL & R. ROJAS, 1996. Catalogue of cuban fossil Elasmobranchii (Paleocene to Pliocene) and paleogeographic implications of their Lower to Middle Miocene occurrence. The Journal of the Geological Society of Jamaica 31: 7-21.
LERICHE, M., 1927. Les poissons de la Molasse suisse. Memoires de la Societé Paleontologique Suisse 46-47: 1-120.
LONGBOTTOM, A. E., 1979. Miocene shark's teeth from Ecuador. Bulletin of the British Museum Natural History - Geology 32(1): 57-70.
MAURY, C. J., 1925. Fósseis terciários do Brasil com descripção de novas formas cretáceas. Monographia do Serviço Geológico e Mineralógico do Brasil IV: 1-665.
NAYLOR, G. J. P & L. F. MARCUS, 1994. Identifying isolated shark teeth of the genus Carcharhinus to species: Relevance for tracking phyletic change through the fossil record. American Museum Novitates 3109: 1-53.
PURDY, R., J. H. MCLELLAN, V. P. SCHNEIDER, S. P. APPLEGATE, R. MEYER & R. SLAUGHTER. 2001. The Neogene sharks, rays and bony fishes from Lee Creek Mine, aurora, North Carolina. Smithsonian Contributions to Paleobiology 90: 71-202.
RAMOS, M. I. F., V. A. TÁVORA, M. P. PINHEIRO & N. B. BAIA, 2004. Microfósseis. In: D. F. ROSSETTI & A. M. GÓES (Eds.): Neógeno da Amazônia Oriental: 93-107. Museu Paraense Emílio Goeldi, Belém.
REIS, M. A. F., 2005. Chondrichthyan Fauna from the Pirabas Formation, Miocene of Northern Brazil, with Comments on Paleobiogeography. Anuário do Instituto de Geociências 28(2): 31-58.
ROSSETTI, D. F. & A. M. GÓES, 2004. Geologia. In: D. F. ROSSETTI & A. M. GÓES (Eds.): Neógeno da Amazônia Oriental: 13-52. Museu Paraense Emílio Goeldi, Belém.
SANTOS, R. S. & H. TRAVASSOS, 1960. Contribuição à Paleontologia do Estado do Pará. Peixes fósseis da Formação Pirabas. Monografia da divisão de Geologia e Mineralogia, Departamento Nacional da Produção Mineral 16: 1-35.
SANTOS, R. S. & M. S. SALGADO, 1971. Contribuição à Paleontologia do Estado do Pará – Novos restos de peixes da Formação Pirabas. Boletim do Museu Paraense Emílio Goeldi, Nova Série, Geologia 16:1-13.
SPRINGER, V. G., 1964. A revision of the carcharhinid shark genera Scoliodon, Loxodon and Rhizoprionodon. Proceedings of the United States National Museum 115(3493): 559-632.
TÁVORA, V. A., 1998. Ocorrência de uma nova espécie de ostracoda na Formação Pirabas (Eomioceno) – estado do Pará. Boletim do Museu Paraense Emílio Goeldi, série Ciências da Terra 10: 3-15.
TOLEDO, P. M. & D. P. DOMNING, 1989. Fóssil Sirenia (Mammalia– Dugongidae) from the Pirabas Formation (Early Miocene), northern Brazil. Boletim do Museu Paraense Emílio Goeldi, série Ciências da Terra 1(2): 119-145.
 Mailing address:
Mailing address:
Museu Paraense Emílio Goeldi
Editor do Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais
Av. Magalhães Barata, 376
São Braz – CEP 66040-170
Caixa Postal 399
Telefone/fax: 55-91- 3249 -1141
E-mail:boletim@museu-goeldi.br
Recebido:30/03/2009
Aprovado:21/12/2009
Responsabilidade editorial: Hilton Tulio Costi
