










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.6 n.2 Belém ago. 2011
Insetos indutores de galhas em florestas de terra firme e em reflorestamentos com espécies nativas na Amazônia Oriental, Pará, Brasil
Gall-inducing insects in terra firme forest and reforested areas in eastern Amazon, Pará, Brazil
Emmanuel Duarte AlmadaI; Geraldo Wilson Afonso FernandesII
IUniversidade Federal de Minas Gerais. Laboratório de Ecologia Evolutiva e Biodiversidade. Belo Horizonte, Minas Gerais, Brasil. Universidade Estadual de Campinas. Núcleo de Estudos e Pesquisas Ambientais. Campinas, São Paulo, Brasil
IIUniversidade Estadual de Campinas. Núcleo de Estudos e Pesquisas Ambientais. Campinas, São Paulo, Brasil
RESUMO
Neste estudo, apresentamos dados sobre a ocorrência e riqueza de insetos galhadores e suas plantas hospedeiras em florestas de terra firme na Amazônia Oriental, em Porto Trombetas, Pará, Brasil. Nesta região, a Mineração Rio do Norte realiza, desde 1981, um projeto de reflorestamento com espécies nativas em áreas de extração de bauxita. Coletas foram realizadas em 2002, nas estações seca e chuvosa. Foram amostradas 36 áreas de reflorestamentos, com idades de plantio variando de mudas recém-plantadas (menos de 1 ano) até 21 anos, e sete áreas de mata primária. Ao todo, foram encontrados 309 morfotipos de galhas em 255 espécies de plantas pertencentes a 45 famílias botânicas. O grupo de galhadores mais frequente foi os Diptera, da família Cecidomyiidae (97%), sendo também encontrados Coleoptera, Hymenoptera e Lepidoptera. As famílias de plantas com maiores riquezas de espécies de galhadores foram Fabaceae (87), Chrysobalanaceae (12), Burseraceae (18), Annonaceae (15), Clusiaceae (15), Euphorbiaceae (9), Melastomatacaeae (13), Malpighiaceae (17), Apocynaceae (7) e Anacardiaceae (11). Essa diversidade local de galhadores pode ser considerada elevada quando comparada a outros levantamentos realizados no cerrado e na Mata Atlântica.
Palavras-chave: Amazônia. Biodiversidade. Galhas de insetos. Mineração.
ABSTRACT
For the first time, data on the occurrence and richness of gall inducing insects and their host plants in a terra firme forest in the Oriental Amazon region, Porto Trombetas, Pará, Brazil are presented. In 1981 the mining company Mineração Rio do Norte, started a restoration project with native species in areas mined for bauxite. Samples of galls were taken in the dry and the wet seasons of 2002; in 36 reforested sites (from recently planted to 21 years) and in seven primary forest sites. Three hundred and nine different morphotypes of insect galls on 255 host plant species belonging to 45 plant families were recorded. The most frequent galling taxa were Diptera of the Cecidomyiidae family (97%), followed by Coleoptera, Hymenoptera, and Lepidoptera. The host plant families most attacked by galling insects were: Fabaceae (87), Chrysobalanaceae (12), Burseraceae (18), Annonaceae (15), Clusiaceae (15), Euphorbiaceae (9), Melastomatacaeae (13), Malpighiaceae (17), Anacardiaceae (11), and Apocynaceae (7). The data indicate a high richness of gall inducing insects when compared to other vegetation types in Brazil (e.g., cerrado and Atlantic rain forest).
Keywords: Amazonia. Biodiversity. Galling insects. Mining.
INTRODUÇÃO
As galhas são hiperplasias e/ou hipertrofias em tecidos vegetais induzidas por insetos, fungos, vírus, bactérias e nematóides (Mani, 1964; Shorthouse et al., 2005). Nas últimas duas décadas, o estudo das galhas trouxe inúmeras contribuições para a ecologia em geral, em especial no que diz respeito à compreensão da evolução das relações parasita-hospedeiro (Price et al., 1998). Insetos galhadores representam uma guilda altamente diversa e importante de herbívoros (Espírito-Santo & Fernandes, 2007) e têm servido como modelo em inúmeros estudos de relações tróficas, padrões globais de distribuição de espécies, além da compreensão dos efeitos da arquitetura e do sexo da planta hospedeira sobre a aptidão de herbívoros (Araújo et al., 2003; Fagundes et al., 2005; Espírito-Santo et al., 2007).
Os padrões globais de distribuição de insetos galhadores têm sido objeto de intensas discussões no meio científico. Price et al. (1998) apontaram um padrão global de distribuição de insetos galhadores em que as latitudes equivalentes entre 23 e 45 graus N ou S apresentariam valores elevados de riqueza desses herbívoros. Essas regiões corresponderiam a habitats xéricos, ou seja, com clima quente, seco e com vegetação esclerófila, nas quais as galhas representariam uma adaptação ao ambiente adverso. Além disso, ambientes xéricos apresentam menores taxas de mortalidade larval dos galhadores devido a inimigos naturais que incluem fungos, parasitóides e predadores (Fernandes & Price, 1988; Fernandes, 1992; Gange et al., 2002). Recentemente, a lista de fatores de mortalidade entre habitats foi ampliada com a inclusão de mecanismos de defesa da planta, como a reação de hipersensibilidade (Fernandes, 1990). Assim, combinando-se o alto grau de esclerofilia dos tecidos e a baixa mortalidade por resistência da planta, predação e parasitismo, os ambientes xéricos abrigariam elevada riqueza de galhadores quando comparados a ambientes mésicos. Esse modelo, baseado nas condições de estresse higrotérmico e nutricional e nas taxas diferenciais de mortalidade entre ambientes (veja Fernandes & Price, 1988, 1991; Fernandes, 1992), tem dominado as discussões em torno dos mecanismos geradores da distribuição biogeográfica de insetos galhadores (Lara & Fernandes, 1996; Price et al., 1998).
No entanto, alguns estudos têm sugerido outros mecanismos para explicar os padrões locais e regionais da distribuição de insetos galhadores. Wright & Samways (1998), estudando a vegetação de Fynbos, na África do Sul, encontraram forte influência da composição florística sobre a riqueza de galhadores. Neste ambiente, geralmente cada espécie de planta abriga uma ou poucas espécies de galhadores, sendo a riqueza de galhadores uma função direta da riqueza de plantas hospedeiras.
Da mesma forma, Blanche & Westoby (1995), estudando as florestas de eucalipto na Austrália, encontraram forte correspondência entre a composição florística e a riqueza de galhadores, medida pela fertilidade do solo. Os resultados encontrados por esses pesquisadores indicaram que a presença de algumas espécies de Eucalyptus é um elemento determinante para a riqueza de galhadores naqueles ambientes. O mesmo mecanismo foi apontado por Fernandes (1992), em um estudo na indonésia, o qual verificou a importância da composição florística como um fator para explicar os padrões de distribuição geográfica de insetos galhadores. No referido estudo, o autor encontrou uma correlação positiva significativa entre o tamanho das famílias botânicas e a riqueza de espécies de galhadores.
Outro mecanismo relevante na regulação da riqueza de insetos galhadores foi apontado por Mendonça (2001), que discute o papel da sincronização da disponibilidade de recursos. Segundo esse autor, o padrão encontrado por Price et al. (1998) pode ser explicado pela sincronicidade na formação de tecidos jovens típica das vegetações mediterrânicas. Nesses ambientes, quando comparados com florestas tropicais chuvosas, tais como a Amazônia, a maior previsibilidade e disponibilidade de sítios de oviposição favoreceriam maior diversificação desses herbívoros.
A baixa riqueza de insetos galhadores esperada para as florestas tropicais úmidas no modelo proposto por Price et al. (1998) pode ser, por outro lado, resultado do pequeno número de estudos dessa guilda de insetos nessas regiões. Até o momento, há apenas uma publicação que faz referência à riqueza de galhas e suas plantas hospedeiras na Amazônia (Julião et al., 2005). Os estudos realizados por Price et al. (1998) e Yukawa et al. (2001) na região amazônica foram pontuais e realizados no sub-bosque de tal formação tropical. Apesar de escassos, tais levantamentos e listas de galhas são fundamentais para o estudo de padrões de diversidade, aliados à necessidade de melhor conhecimento taxonômico das espécies indutoras de galhas e suas respectivas plantas hospedeiras nesta floresta tropical. Por outro lado, a maioria dos esforços amostrais no Brasil se concentrou em áreas de cerrado, em especial de campos rupestres (Fernandes et al., 1997, 2005; Lara & Fernandes, 1996; Gonçalves-Alvim & Fernandes, 2001; Lara et al., 2002).
Neste trabalho, são apresentados dados de morfotipos de galhas induzidas por insetos e suas respectivas plantas hospedeiras em florestas de terra firme da Amazônia Oriental.
MATERIAL E MÉTODOS
Este estudo foi conduzido em áreas de reflorestamento e florestas primárias de terra firme, em Porto Trombetas, localizado na Floresta Nacional Saracá Taquera, com uma altitude de 180 m, 65 km a nordeste da cidade de Oriximiná e a 30 km do rio Trombetas, no estado do Pará, Brasil (1o 40' S; 56o 27' W). A média anual de precipitação em Porto Trombetas (1970-1994) é de 2.185 ± 964 (S.E.) mm, com estações chuvosas (inverno) e seca (verão), distintas médias mensais de precipitação excedendo 10 mm em todos os meses, exceto no período de julho a outubro. A média máxima e mínima de temperatura é de 34.6 e 19.9 oC, respectivamente. Os solos nas áreas de extração de bauxita são latosolos argilosos amarelos e ácidos, com uma fina camada de húmus (Ferraz, 1993). A vegetação regional é formada por uma floresta equatorial perene, com dossel chegando a uma altura de 20 a 35 m, com árvores emergentes de até 45 m de altura (Knowles & Parrotta, 1995, 1997). A floresta, na região da mineração, era, até recentemente, largamente inacessível e sem distúrbios por derrubada ou retirada de madeira por mais de 200 anos. A restauração florestal ocorreu inicialmente com uma combinação de espécies nativas de diferentes estágios sucessionais, sendo que seu progresso está associado ao recrutamento do banco de sementes, bem como a processos de dispersão zoocórica e anemocórica (Parrota & Knowles, 1999, 2001; Salomão et al., 2007).
A amostragem de galhas ocorreu durante 20 dias consecutivos na estação seca (julho) e 20 dias consecutivos na chuvosa (dezembro) de 2002 (n = 40 dias de coleta). Amostragens foram realizadas em áreas restauradas, correspondendo aos seguintes anos de plantio: 1981 (n = 2), 1982 (n = 2), 1983 (n = 1), 1984 (n = 2), 1985 (n = 2), 1986 (n = 2), 1987 (n = 2), 1988 (n = 1), 1992 (n = 1), 1993 (n = 2), 1994 (n = 2), 1995 (n = 2), 1996 (n = 2), 1997 (n = 2), 1998 (n = 3), 1999 (n = 2), 2000 (n = 2), 2001 (n = 2), 2002 (n = 2). Assim, foram coletadas galhas de insetos em 36 áreas reflorestadas com espécies nativas da Amazônia, variando de áreas com mudas recém-plantadas até as que apresentam 21 anos de idade de plantio, quando realizado o estudo. A distância mínima entre as áreas foi de 500 m (veja Fernandes & Price, 1988).
Em cada área de reflorestamento foram realizadas três amostragens de uma hora por amostragem (totalizando 6 horas/área amostrada = 216 h). Sete áreas de mata primária também foram amostradas utilizando-se o mesmo protocolo (42 h). A cada amostragem, durante caminhadas de 1 h, todas as plantas com até três metros de altura foram vistoriadas quanto à presença de galhas.
Foi coletado material suficiente das plantas atacadas por galhas para identificá-las e caracterizar o inseto galhador. Para detalhes da metodologia de campo, veja Price et al. (1998). Em laboratório, os insetos indutores foram identificados até o nível taxonômico de ordem ou família, sempre que possível, sendo também realizada a descrição dos principais aspectos morfológicos das galhas, tais como cor, forma, órgão atacado, número de lojas e presença de tricomas (Stone & Schönrogge, 2003). Sempre que possível, as plantas hospedeiras foram identificadas por especialistas até o nível de espécie.
Morfotipos de galhas foram utilizados como um indicador de espécies de galhadores porque praticamente todas as espécies de insetos galhadores amazônicos são novas para a ciência. No entanto, o uso de morfotipos de galhas, associado ao órgão de ocorrência, é aceitável como substituto para espécies de insetos galhadores (Floate et al., 1996; Price et al., 1998; Cuevas-Reyes et al., 2004; Carneiro et al., 2009), dada sua morfologia única e alta especificidade quanto à planta hospedeira e ao órgão da planta atacado (Dreger-Jauffret & Shorthouse, 1992; Floate et al., 1996; Shorthouse et al., 2005)1.
RESULTADOS
A riqueza de insetos galhadores foi elevada, sendo encontradas 309 morfoespécies de insetos galhadores em 255 espécies de plantas hospedeiras, com uma média de 1,2 espécies galhadoras por planta hospedeira (Apêndice). Cento e sessenta e sete morfotipos de galhas ocorrem nas áreas de mata primária, sendo que destas, 137 ocorrem exclusivamente nesses ambientes e, dos 190 morfotipos de galhas que ocorrem nos reflorestamentos, 169 são exclusivos destas áreas. Assim, apenas 22 morfotipos de galhas, em 19 espécies de plantas hospedeiras, são compartilhados pelas áreas de mata primária e as áreas de reflorestamento.
As famílias de plantas com maiores riquezas de espécies de galhadores foram Fabaceae (87), Chrysobalanaceae (12), Burseraceae (18), Annonaceae (15), Clusiaceae (15), Euphorbiaceae (9), Melastomatacaeae (13), Malpighiaceae (17), Apocynaceae (7) e Anacardiaceae (11). Das 44 famílias encontradas, 11 apresentaram apenas uma espécie de galhador. De todas as espécies de hospedeiras encontradas, apenas 11% eram lianas e 5% arbustos, mostrando que a maior parte da comunidade de galhadores concentra-se nas espécies de hábito arbóreo. Infelizmente, não foi possível identificar parte das plantas hospedeiras até o nível específico, visto que praticamente todo material coletado para confecção de exsicatas proveio de indivíduos em estado não fértil.
Do total de morfotipos de galhas encontrados, 67% são verdes, 48% são discóides, 89% ocorrem nas folhas e 96% são glabras. O grupo de galhadores mais frequente foi o Cecidomyiidae (301), sendo também encontrados insetos das ordens Coleoptera (5), Hymenoptera (1), Homoptera (1) e Lepidoptera (1) (Figuras 1-276). Os galhadores de inga (20) e Protium (16) representam mais de 10% das morfoespécies de galha e Bellucia (2), Goupia (3) e Vismia (10), em torno de 5%.
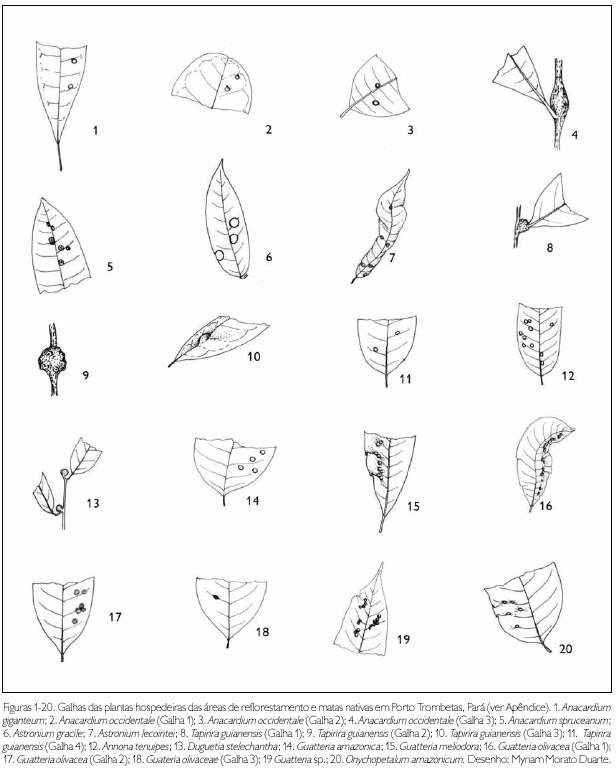

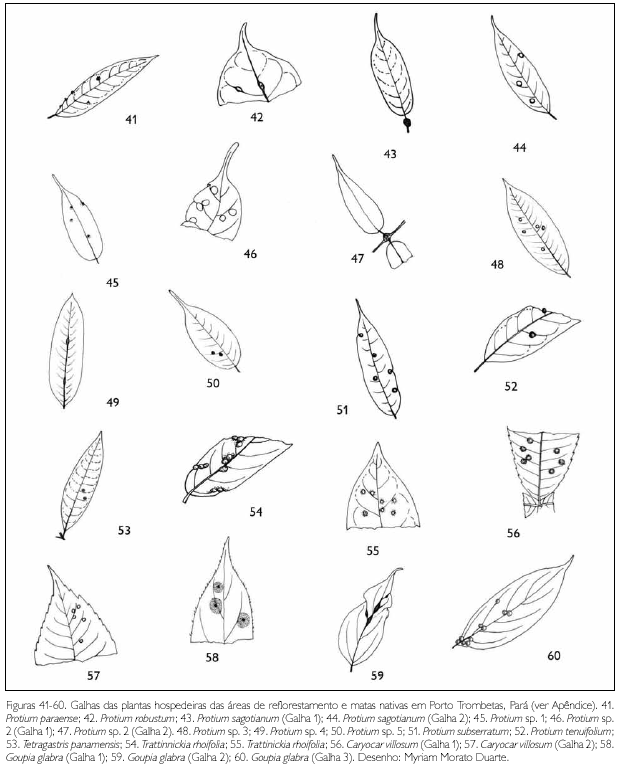
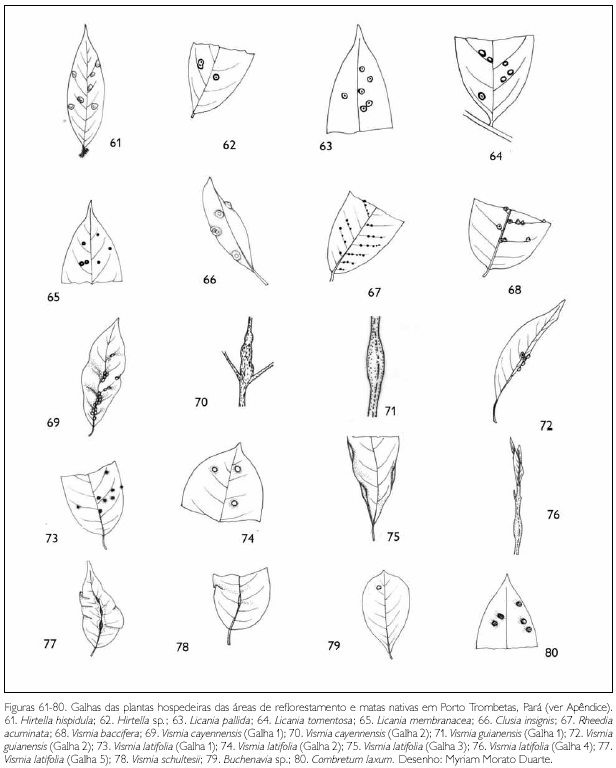
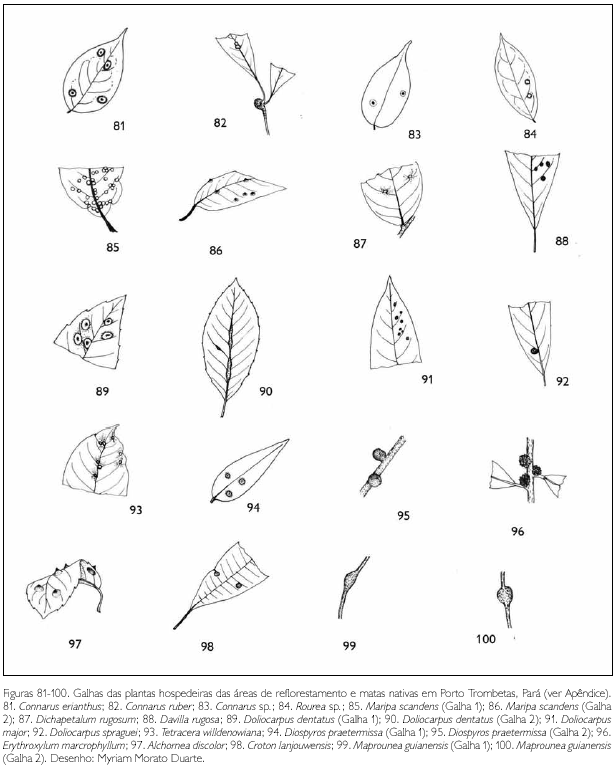
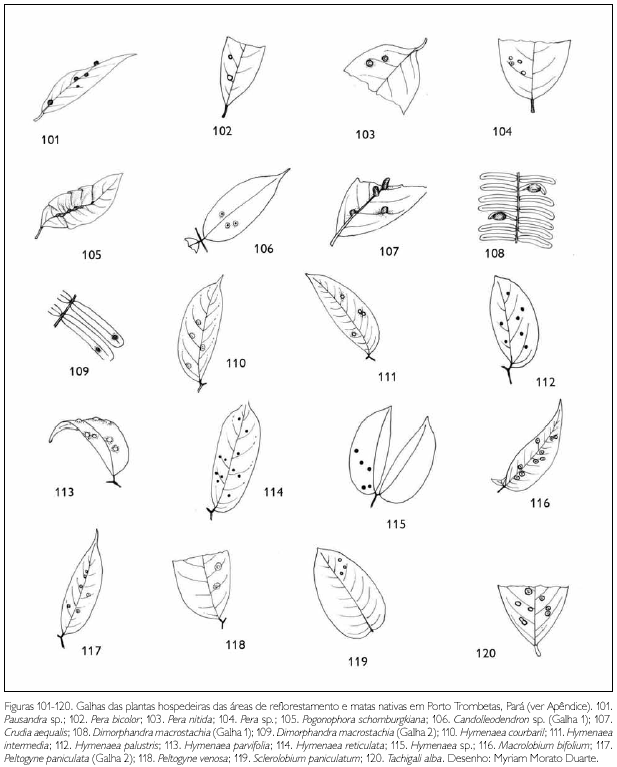
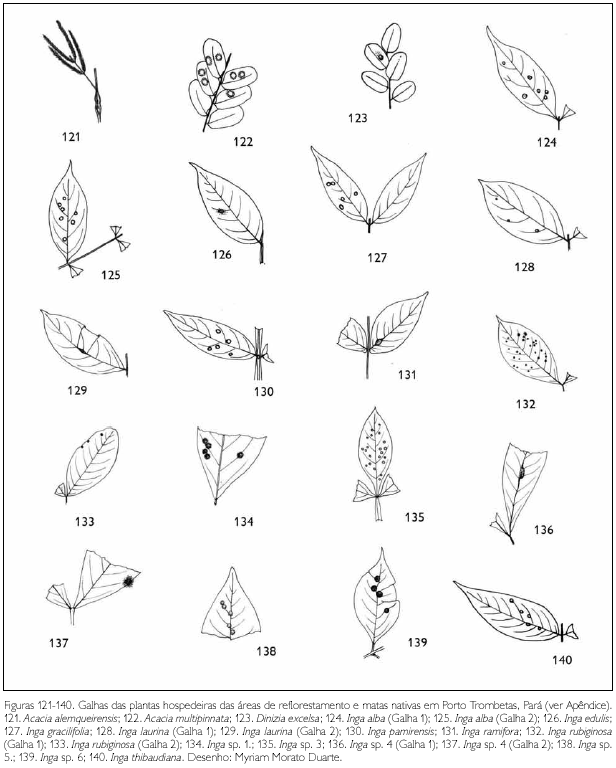
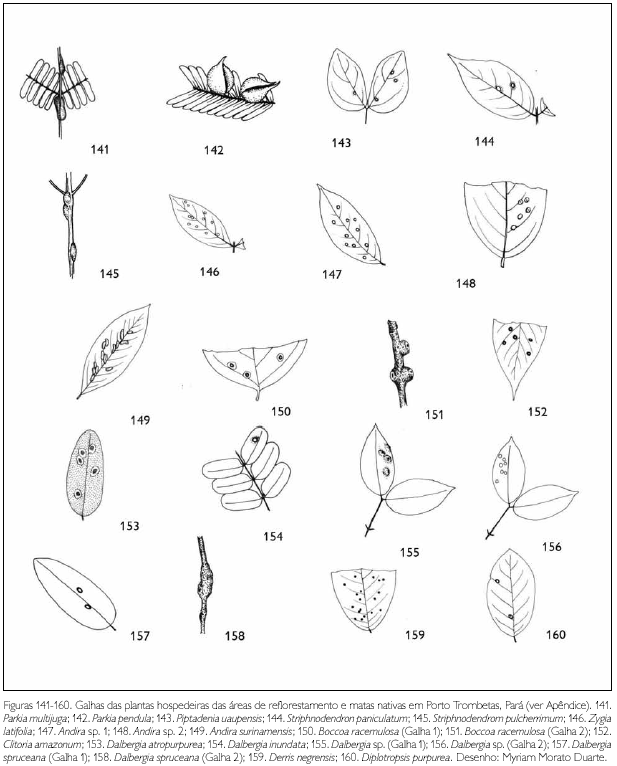

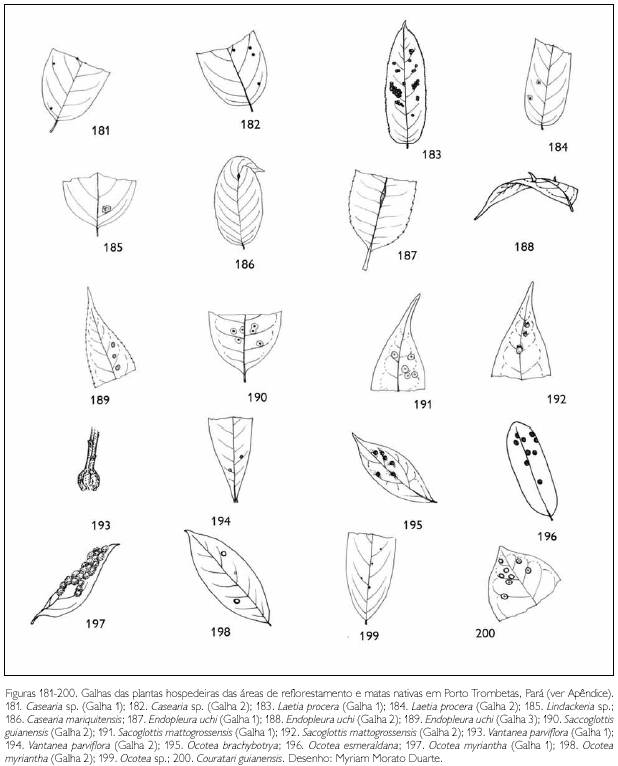

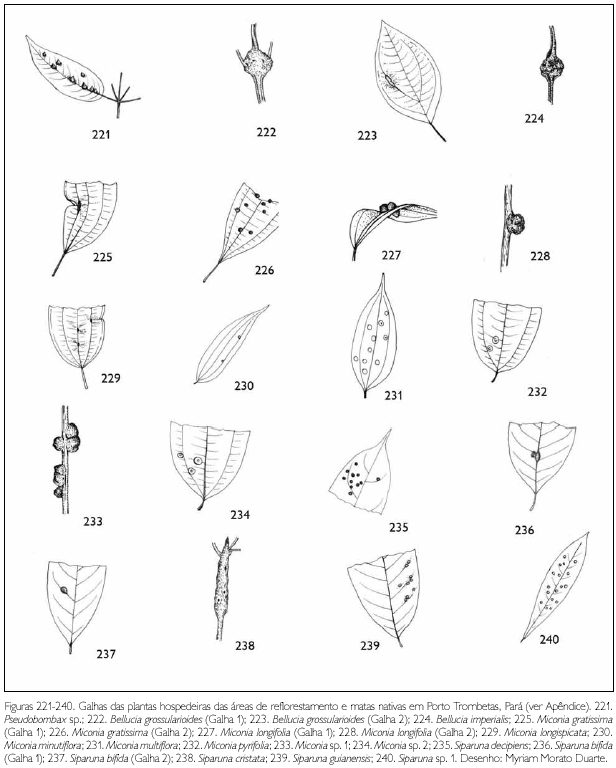

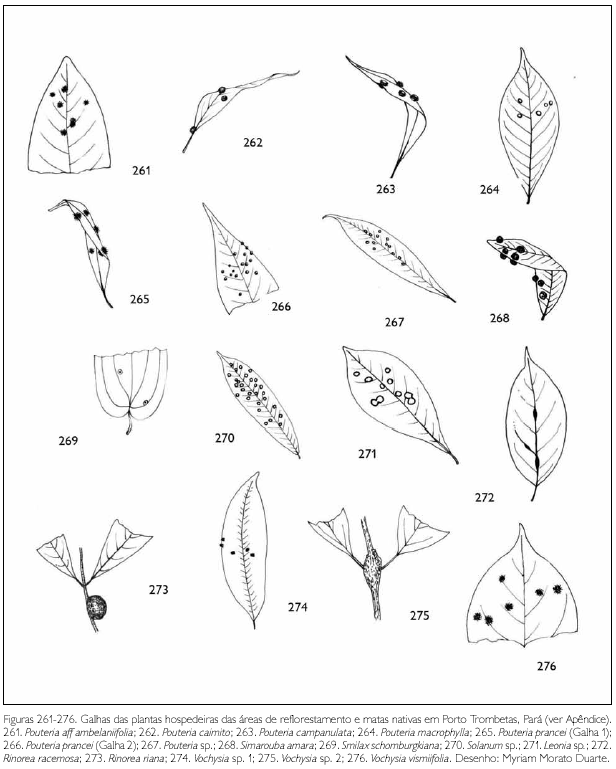
DISCUSSÃO
A riqueza de insetos galhadores deste estudo (309) foi elevada, quando comparada a outros dados previamente encontrados para a floresta amazônica. Em um estudo com insetos galhadores próximo a Manaus, Yukawa et al. (2001) encontraram 84 espécies, enquanto Julião (2007) encontrou valores similares ao do presente estudo para a riqueza de galhadores de dossel em florestas de terra firme próximas a Manaus (303 morfoespécies). Fernandes et al. (1997) encontraram 236 morfoespécies de galhadores no Vale do Jequitinhonha e Maia & Fernandes (2004) descreveram 137 morfoespécies na Serra de São José, Minas Gerais. Ainda que se considerem as diferenças de esforço amostral entre os trabalhos citados e a riqueza encontrada no presente trabalho, o presente estudo sugere que as florestas de terra firme da Amazônia podem ser uma das áreas com maior diversidade de insetos galhadores.
Os resultados do presente trabalho também reforçam a importância da família Cecidomyiidae como o principal grupo taxonômico entre os galhadores na região neotropical, respondendo por mais de 90% das morfoespécies de galhadores (Mani, 1964; Carneiro et al., 2009).
A composição das principais famílias de plantas hospedeiras também foi distinta da encontrada em outros estudos. Em remanescentes de Mata Atlântica, em Minas Gerais, as famílias com maior número de insetos galhadores foram Asteraceae, Myrtaceae, Bignoniaceae e Melastomataceae (Fernandes et al., 2002). É interessante ressaltar a contribuição de Melastomataceae para a riqueza de insetos galhadores neste estudo, já que é uma das principais famílias que compõem o sub-bosque das florestas de terra firme na Amazônia (Ribeiro et al., 1999). No levantamento de galhas feito por Julião et al. (2005) em áreas de várzea amazônica em Mamirauá, Melastomataceae não aparece entre as principais famílias de plantas hospedeiras. Já nos estudos realizados em capões de mata no Pantanal sul-matogrossense, nenhum galhador foi encontrado em Melastomataceae, sendo que Bignoniaceae foi a principal família hospedeira, seguida de Fabaceae, Sapindaceae e Hippocrantaceae (Julião et al., 2004).
De maneira geral, a comunidade vegetal na Amazônia não ocorre sob condições de estresse hídrico, a não ser nas áreas inundáveis de várzea e igapó, onde, durante as épocas de cheia, as raízes das árvores ficam sob condições de baixas concentrações de oxigênio, limitando as taxas metabólicas de toda a comunidade vegetal (Schongart et al., 2005). Nas florestas de terra firme, que representam 85% da área amazônica (Ribeiro et al., 1999), a disponibilidade de água no solo é elevada durante praticamente todo o ano. No sub-bosque da floresta, a umidade mantém-se elevada e não há grande incidência de ventos e radiação. Assim, é provável que no sub-bosque das florestas de terra firme outros fatores ambientais (tais como a abundância das espécies hospedeiras, riqueza de plantas ou ataque por inimigos naturais), que não o estresse higrotérmico, sejam os principais determinantes para a riqueza de insetos galhadores.
No entanto, em particular, o dossel das florestas pluviais, a despeito das altas taxas de precipitação, apresenta condições de forte estresse devido à menor umidade do ar e à maior velocidade dos ventos, representando um cenário cujas pressões ambientais se assemelham às do modelo proposto por Fernandes & Price (1988), onde o estresse higrotérmico e nutricional, além da esclerofilia, são fatores-chave para a riqueza de insetos galhadores. Julião (2007), estudando o efeito de variações locais da fertilidade do solo sobre a riqueza de galhas de dossel na Amazônia, encontrou correlação negativa entre a disponibilidade de nutrientes e a riqueza de galhadores em florestas de terra firme próximas a Manaus. No estudo de caso aqui apresentado, as áreas de reflorestamento, com elevada taxa de radiação e transpiração, mais próximas das condições do ambiente de dossel, também apresentaram maior riqueza de insetos galhadores. Além disso, muitas espécies vegetais encontradas no sub-bosque e nas áreas de reflorestamento, como Parkiapendula e Minquartiaguianensis, na verdade, são indivíduos jovens de árvores que compõem o dossel da floresta. Apesar das previsões de baixa diversidade de insetos galhadores para a região amazônica (Price et al., 1998), este estudo indica que as florestas de terra firme podem representar uma das regiões biogeográficas mais ricas em relação a essa guilda de herbívoros. Julião et al. (2005), Julião (2007) e Ribeiro & Basset (2007) apontam o dossel da floresta como um ambiente rico em galhadores, dado seu maior grau de esclerofilia, bem como de condições de estresse higrotérmico.
Como apontado por alguns autores (por exemplo, Fernandes, 1992; Fernandes & Price, 1988), a presença de super-hospedeiros pode ser um elemento importante para determinar a composição da comunidade de galhadores. No presente estudo, apenas os gêneros Inga e Protium responderam por aproximadamente 10% dos morfotipos de galhas encontradas. Algo similar pode ser encontrado nas áreas de cerrado, onde o gênero Baccharis é um super-hospedeiro (Fernandes et al., 1996; Espírito Santo et al., 2007), ou ainda em relação ao gênero Eucalyptus na Austrália (Blanche & Westoby, 1995).
CONCLUSÃO
Os dados indicam uma elevada riqueza de insetos galhadores para a região amazônica e, ainda, que o sub-bosque pode ter um papel relevante neste contexto. Todavia, evidencia a necessidade de mais estudos comparando os estratos desta floresta tropical chuvosa, levando-se em consideração as condições ambientais contrastantes entre o dossel e o sub-bosque da floresta para a história evolutiva desses herbívoros.
Corroborando os resultados de outros estudos, este trabalho também aponta a família Cecidomyiidae como o principal grupo entre os indutores de galhas em região neotropical, com menor contribuição de outros grupos de insetos, como Hymenoptera e Lepidoptera.
Apesar da elevada riqueza de insetos galhadores encontrada neste estudo, a floresta amazônica continua a ser o bioma com menor esforço amostral para este grupo de herbívoros. A maioria dos estudos evolutivos e ecológicos de insetos galhadores foi realizada em zonas temperadas ou em áreas mais áridas da região tropical. Esta lacuna do conhecimento e os resultados aqui apresentados indicam a necessidade de estudos mais detalhados para compreender o significado adaptativo das galhas, bem como as forças evolutivas que atuam sobre os insetos galhadores nas florestas tropicais úmidas.
AGRADECIMENTOS
Agradecemos a Y Oki, R. Isaias, M. M. Espírito-Santo, M. A. A. Carneiro e a dois revisores anônimos pela revisão e sugestões neste manuscrito. Agradecemos à equipe da Floresta Nacional Saracá Taquera e do Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA), à Mineração Rio do Norte e à Planta Tecnologia Ambiental pelo apoio logístico; e ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (30.9633/2007-9) pelas bolsas concedidas. Este artigo é parte da dissertação de mestrado de E. D. Almada, apresentada ao Programa de Pós-graduação em Ecologia, Conservação e Manejo da Vida Silvestre Instituto de Ciências Biológicas/Universidade Federal de Minas Gerais (ICB/UFMG).
REFERÊNCIAS
ARAÚJO, A. P. A., G. W. FERNANDES & M. A. A. CARNEIRO, 2003. Efeitos do sexo, do vigor e do tamanho da planta hospedeira sobre a distribuição de insetos indutores de galhas em Baccharis camporum DC (Asteraceae). Revista Brasileira de Entomologia 47(4): 483-490.
BLANCHE, K. R. & M. WESTOBY, 1995. Gall-forming insect diversity is linked to soil fertility via host plant taxon. Ecology 76(7): 2334-2337.
CARNEIRO, M. A. A., C. S. A. BRANCO, C. E. D. BRAGA, E. D. ALMADA, M. B. M. COSTA, G. W. FERNANDES & V. C. MAIA, 2009. Are gall midge species (Diptera: Cecidomyiidae) host plant specialists? Revista Brasileira de Entomologia 53(3): 365-378.
CUEVAS-REYES, P., M. QUESADA, P. HANSON, R. DIRZO & K. OYAMA, 2004. Diversity of gall-inducing insects in a Mexican tropical dry forest: the importance of plant species richness, life forms, host plant age and plant density. Journal of Ecology 92(4): 707-716.
DREGER-JAUFFRET, F. & J. D. SHORTHOUSE, 1992. Diversity of gall-inducing insects and their galls. In: J. D. SHORTHOUSE & O. ROHFRITSCH (Eds.): Biology of Insect-Induced Galls: 8-33. Oxford University Press, Oxford.
ESPÍRITO-SANTO, M. M. & G. W. FERNANDES, 2007. How many species of Gall-Inducing insects are there on earth, and where are they? Annals of the Entomological Society of America 100(2): 95-99.
ESPÍRITO-SANTO, M. M., F. S. NEVES, F. R. ANDRADE-NETO & G. W. FERNANDES, 2007. Plant architecture and meristem dynamics as the mechanisms determining the diversity of gall-inducing insects. Oecologia 153(2): 353-364.
FAGUNDES, M., F. NEVES & G. W. FERNANDES, 2005. Direct and indirect interactions involving ants, insect herbivores, parasitoids and the host plant Baccharis dracunculifolia (Asteraceae). Ecological Entomology 30(1): 28-35.
FERNANDES, G. W., 1990. Hypersensitivity: a neglected plant resistance mechanism against insect herbivores. Environmental Entomology 19(5): 1173-1182.
FERNANDES, G. W., 1992. Plant family size and age effects on insular gall-forming species richness. Global Ecology and Biogeography Letters 2(3): 71-74.
FERNANDES, G. W. & P. W. PRICE, 1988. Biogeographical gradients in galling species richness. Oecologia 76(2): 161-167.
FERNANDES, G. W. & P. W. PRICE, 1991. Comparisons of tropical and temperate galling species richness: the roles of environmental harshness and plant nutrient status. In: P. W. PRICE, T. M. LEWISON, G. W. FERNANDES & W. BENSON (Eds.): Plant-animal interactions: evolutionary in tropical and temperate regions: 91-115. Wiley, NewYork.
FERNANDES, G. W., M. A. A. CARNEIRO, A. C. F. LARA, L. A. ALLAIN, G. I. ANDRADE, G. R. JULIÃO, T. C. REIS & I. M. SILVA, 1996. Galling insects on neotropical species of Baccharis (Asteraceae). Tropical Zoology 9(2): 315-332.
FERNANDES, G. W., R. C. ARAÚJO, S. C. ARAÚJO, J. A. LOMBARDI, A. S. PAULA, R. LOYOLA & T. G. CORNELISSEN, 1997. Insect Galls From Jequitinhonha Valley, Minas Gerais, Brazil. Naturalia 22(3): 221-224.
FERNANDES, G. W., G. R. JULIÃO, R. C. ARAÚJO, S. C. ARAÚJO, J. A. LOMBARDI, D. NEGREIROS & M. A. A. CARNEIRO, 2002. Distribution and morphology of insect galls of the Rio Doce Valley, Brasil. Naturalia 26(3): 211-244.
FERNANDES, G. W., S. J. GONÇALVES-ALVIM & M. A. A. CARNEIRO, 2005. Habitat-driven effects on the diversity of gall-inducing insects in the Brazilian Cerrado Biology, ecology and evolution of gall-inducing arthropods. In: A. RAMAN, C. W. SCHAEFER & T. M. WITHERS (Orgs.): Biology, Ecology, and evolution of gall-inducing arthropods: 693-708. Scince Publishers Inc., Enfield.
FERRAZ, J. B. S., 1993. Soil factors influencing the reforestation on mining sites in Amazonia. In: H. LIETH & M. LOHRANN (Eds.): Restoration of tropical forest ecosystems: 47-52. Kluwer Academic, Dordrecht.
FLOATE, K. D., G. W. FERNANDES & J. A. NILSSON, 1996. Distinguishing intrapopulational categories of plants by their insect faunas: galls on rabbitbrush. Oecologia 105(2): 221-229.
GANGE, A. C., R. CROFT & W. P. WU, 2002. Gall insect and endophytic fungal co-occurrence in a xeric and mesic site. Ecological Entomology 27(3): 362-365.
GONÇALVES-ALVIM, S. J. & G. W. FERNANDES, 2001. Comunidades de insetos galhadores (Insecta) em diferentes fisionomias do cerrado em Minas Gerais, Brasil. Revista Brasileira de Zoologia 18(1): 289-305.
JULIÃO, G. R., 2007. Riqueza e abundância de insetos galhadores associados ao dossel de florestas de terra firme, várzea e igapó da Amazônia Central: 1-158. Tese (Doutorado em Ecologia) - Instituto Nacional de Pesquisas da Amazônia, Manaus, Amazonas. Disponível em: <http://tede.inpa.gov.br/tde_arquivos/2/TDE-2008-12-26T142824Z-116/Publico/Tese_Genimar_Pt_1.pdf>. Acesso em: 20 julho 2011.
JULIÃO, G. R., M. E. C. AMARAL, G. W. FERNANDES & E. G. OLIVEIRA, 2004. Edge effect and species-area relationships in the gall-forming insect fauna of natural forest patches in the Brazilian Pantanal. Biodiversity and Conservation 13(11): 2055-2066.
JULIÃO, G. R., E. M. VENTICINQUE & G. W. FERNANDES, 2005. Richness and abundance of gall-forming insects in the Mamirauá Varzea, a flooded Amazonian forest. Uakari 1(1): 39-42.
KNOWLES, O. H. & J. A. PARROTTA, 1995. Amazonian forest restoration: an innovative system for native species selection based on phenological data and performance indices. Commonwealth Forestry Review 74(3): 230-243.
KNOWLES, O. H. & J. A. PARROTTA, 1997. Phenological observations and tree seed characteristics in an equatorial moist forest at Trombetas, Pará State, Brazil. In: H. LIETH & M. D. SCHWARTZ (Eds.): Phenology in Seasonal Climates I: 67-84. Backhuys, Leiden.
LARA, A. C. F. & G. W. FERNANDES, 1996. The highest diversity of galling insects: Serra do Cipó, Brazil. Biodiversity Letters 3(3): 111-114.
LARA, A. C. F., G. W. FERNANDES & S. J. GONÇALVES-ALVIM, 2002. Tests of hypotheses on patterns of gall distribution along an altitudinal gradient. Tropical Zoology 15(2): 219-232.
MAIA, V. C. & G. W. FERNANDES, 2004. Insect galls from Serra de São José (Tiradentes, MG, Brazil). Brazilian Journal of Biology 6(3): 423-445.
MANI, M. S., 1964. Ecology of plant galls: 1-434. W. Junk Publisher, Netherlands.
MENDONÇA, M. D., 2001. Galling insect diversity patterns: the resource synchronization hypothesis. Oikos 95(1): 171-176.
PARROTTA, J. A. & O. H. KNOWLES, 1999. Restoration of tropical moist forests on bauxite-mined lands in the Brazilian Amazon. Restoration Ecology 7(2): 103-116.
PARROTTA, J. A. & O. H. KNOWLES, 2001. Restoring tropical forests on lands mined for bauxite: examples from the Brazilian Amazon. Ecological Engineering 17(2-3): 219-239.
PRICE, P. W., G. W. FERNANDES, A. C. LARA, J. BRAN, H. BARRIOS, M. G. RIGHT, S. P. RIBEIRO & N. ROTHCLIFF, 1998. Global patterns in local number of insect galling species. Journal of Biogeography 25(3): 581-591.
RIBEIRO, J. E. L. S., M. J. G. HOPKINS, A. VICENTINI, C. A. SOTHERS, M. A. S. COSTA, J. M. BRITOM, M. A. D. SOUZA, L. H. P. MARTINS, L. G. LOHMANN, P. A. C. L. ASSUNÇÃO, E. C. PEREIRA, C. F. SILVA, M. R. MESQUITA & L. C. PROCÓPIO, 1999. Flora da Reserva Ducke. Guia de identificação das plantas vasculares de uma floresta de terra-firme na Amazonia Central: 1-800. INPA-DFID, Manaus.
RIBEIRO, S. P. & Y. BASSET 2007. Gall-forming and free-feeding herbivory along vertical gradients in a lowland tropical rainforest: the importance of leaf sclerophylly. Ecography 30(5): 663-672.
SALOMÃO, R. P., N. A. ROSA & K. A. C. MORAIS, 2007. Dinâmica da regeneração natural de árvores em áreas mineradas na Amazônia. Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais 2(2): 85-139.
SCHONGART, J., M. T. F. PIEDADE & F. WITTMANN, 2005. Wood growth patterns of Macrolobium acaciifolium (Benth.) Benth. (Fabaceae) in Amazonian black-water and white-water floodplain forests. Oecologia 145(3): 454-461.
SHORTHOUSE, J. D., D. WOOL & A. RAMAN, 2005. Gall-inducing insects - nature's most sophisticated herbivores. Basic and Applied Ecology 6(5): 407-411.
STONE, G. N. & K. SCHÖNROGGE, 2003. The adaptive significance of insect gall morphology. Trends in Ecology and Evolution 18(10): 512-522.
YUKAWA, J., M. TOKUDA, N. UECHI & S. SATO, 2001. Species richness of galling arthropods in Manaus, Amazon and the surroundings of the Iguassu falls. Esakia 41(1): 11-15.
WRIGHT, M. G. & M. J. SAMWAYS, 1998. Insect species richness tracking plant species richness in a diverse flora: gall-insects in the Cape floristic Region, South Africa. Oecologia 115(3): 427-433.
 Autor para correspondência:
Autor para correspondência:
Emmanuel Duarte Almada.
Universidade Federal de Minas Gerais.
Instituto de Ciências Biológicas.
Avenida Presidente Antonio Carlos, 6627.
Campus Pampulha. Belo Horizonte, MG, Brasil.
CEP 30161-970
(almadaceae@gmail.com).
Recebido em 09/03/2010
Aprovado em 29/06/2011
Responsabilidade editorial: Marinus Hoogmoed
1Para uma revisão mais detalhada sobre o uso de morfotipos de galhas na avaliação da riqueza de espécies de galhadores, veja Carneiro et al. (2009).
