










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.6 n.3 Belém dez. 2011
Similaridade de espécies arbóreas em função da distância em uma floresta ombrófila na Floresta Nacional de Saracá-Taquera, Pará
Similarity of tree species in relation to distance in a rain forest in the National Forest Saracá-Taquera, Pará
Leandro Valle FerreiraI; Rafael de Paiva SalomãoII; Darley Calderaro Leal MatosI; Jorge Luis Gavina PereiraIII
IMuseu Paraense Emílio Goeldi. Coordenação de Ciências da Terra e Ecologia. Belém, Pará, Brasil
IIMuseu Paraense Emílio Goeldi. Coordenação de Botânica. Belém, Pará, Brasil
IIIMuseu Paraense Emílio Goeldi. Coordenação de Pesquisa e Pós-Graduação. Belém, Pará, Brasil
RESUMO
A distribuição de espécies é uma questão complexa. Atualmente, existem dois principais modelos usados para explicar essa distribuição: a Teoria da Neutralidade e a Teoria de Nicho. A primeira prediz que a coexistência de espécies é resultado de um equilíbrio entre imigração e extinção, enquanto a segunda prediz que os recursos naturais são utilizados diferencialmente entre as espécies. Estudos realizados em regiões tropicais têm demonstrado que a similaridade de espécies diminui com a distância geográfica. Contudo, poucos estudos foram realizados comparando a similaridade de espécies em relação à distância geográfica, em escala local, onde outras variáveis, como topografia, altitude e tipo de solo não variam. O objetivo deste estudo é testar se existem diferenças na similaridade de espécies em relação à distância geográfica entre parcelas, em um platô de 1.500 hectares recoberto por uma floresta ombrófila densa, na Floresta Nacional Sacará-Taquera, estado do Pará. Neste local, foram distribuídas 179 parcelas de 10 x 250 metros, cobrindo a superfície total do platô. A distância entre as parcelas variou de 200 metros a 9 quilômetros. Nas 179 parcelas amostradas foram identificadas 631 espécies. A curva cumulativa de espécies apresentou uma nítida tendência assíntota. As estimativas de riqueza de espécies, utilizando os estimadores de riqueza Jackknife 1 e 2, foram de 720 e 733 espécies, respectivamente. A maioria das espécies apresentou baixa frequência absoluta nas parcelas amostradas, um padrão típico para a floresta amazônica; do total de 631 espécies, 442 (70% do total) ocorreram em menos de 10% do total das parcelas. Houve uma correlação negativa entre a similaridade de espécies e a distância geográfica das parcelas no platô. Os menores índices de similaridade de Sórensen, variando de 13% a 19%, foram obtidos entre parcelas a distâncias médias de nove quilômetros entre si, enquanto os maiores índices de similaridade, variando de 50% a 62%, foram obtidos a distâncias menores do que um quilômetro entre si. Este estudo corrobora parcialmente a Teoria da Neutralidade, na qual a similaridade entre espécies diminui com a distância geográfica. Contudo, o valor da correlação obtida entre similaridade e distância geográfica (r = -0.44) indica que outros fatores influenciam esta diferenciação, mostrando que a similaridade de espécies em regiões tropicais pode ser explicada por outros fatores, biótica e abiótica, além da distância.
Palavras-chave: Amazônia. Distância. Espécies raras. Similaridade de espécies.
ABSTRACT
The distribution of species in tropical regions is very complex. Two principal models are commonly employed to explain species distributions: the Neutral Theory, and the Niche Theory. The former predicts that species coexistence is the result of a balance between immigration and extinction, while the Niche Theory predicts that natural resources are used differently among species. Studies carried out in tropical regions have demonstrated that species similarity decreases with geographical distance. However, few studies have been done comparing changes in species similarity with geographical distance on a local scale, where physical variables such as topography, altitude and soil type do not vary. The objective of this study is to test differences in species similarity with geographical distance from botanical plots on an Amazonian plateau in the Sacará-Taquera National Forest Reserve, in Pará, Brazil. 179 tree plots (10 x 250 m) were systematically distributed across the entire extent of the plateau, with a distance between plots varying between 200 meters and 9 km. A total of 631 species of tree were identified across all plots. The species area curve showed a clear tendency to asymptote. The estimated species richness (using first and second order Jackknife estimators) suggested 720 and 733 species respectively. The majority of species were present in low frequencies in the sampled plots, a pattern typical for the Amazon region. From the total of 631 species, 442 (70%) occurred in less than 10% of all parcels. There was a negative correlation between species similarity and plot distances. The lowest Sórensen similarity measures (ranging from 13% to 16%) were obtained from plots over 6 km apart, while the highest levels of similarity (ranging from 55% to 62%) were found comparing plots less than one kilometer apart. In conclusion, this study partially corroborates the Neutral Theory, where the similarity of species decreases with increasing geographical distances. However, the medium strength correlation between similarity and distance (r = -0.44) indicates that other factors are also important, and showing that the similarity of species in the tropical rain forests may be partly explained by other biotic and abiotic factors, as well as geographical distance.
Keywords: Amazon. Distance. Rare species. Similarity of species.
INTRODUÇÃO
A variação da riqueza e composição de espécies em regiões tropicais têm sido atribuídas a fatores físicos e biológicos, sendo estes a base para os modelos teóricos em ecologia de comunidades (Wright et al., 1993), tais como a Teoria da Neutralidade, de Hubbell (2001), e a Teoria do Nicho (Wright, 2002; Kneitel & Chase, 2004), que propõem explicar os mecanismos condutores da distribuição de espécies em comunidades.
A Teoria da Neutralidade, de Hubbell (2001), assume que todos os indivíduos de uma comunidade são funcionalmente equivalentes e apresentam a mesma probabilidade de migrar, reproduzir-se e morrer. Explica que a coexistência de espécies, em diferentes escalas, é resultado do equilíbrio entre imigração e extinção, em escala local, e entre as taxas de especiação e extinção, em escala regional. Também prediz que a similaridade de espécies em uma comunidade diminui com o aumento da distância geográfica entre locais, independente das diferenças ambientais, sendo esta redução o resultado da limitação de dispersão no espaço. Assim, os mecanismos que geram diferenças nos padrões de composição das espécies estão ligados à capacidade de dispersão dos indivíduos (Hubbell, 2006).
A Teoria do Nicho (Macarthur & Levins, 1964) prediz que os recursos são utilizados diferencialmente entre as espécies, onde a habilidade de uma função se faz à custa de outra, ou seja, a especialização na obtenção de um recurso normalmente vem acompanhada de uma diminuição na eficiência em relação a outro (Condit et al., 2002; Tilman, 2004). Apenas espécies com nichos suficientemente diferenciados podem coexistir na mesma comunidade (Kneitel & Chase, 2004). Dessa forma, o padrão de composição das espécies deve ser determinado por características ambientais que relacionem as espécies capazes de se estabelecer em certo local.
Diversos estudos mostram que a distância geográfica influencia na distribuição de plantas em regiões tropicais (Scudeller et al., 2001; Condit et al., 2002; Ruokolainen & Tuomisto, 2002; Carneiro & Valeriano, 2003), pois a similaridade de espécies diminui com o aumento da distância geográfica entre parcelas, corroborando o que afirma a Teoria da Neutralidade.
A Teoria da Neutralidade tende a explicar adequadamente os padrões de composição de organismos residentes, como plantas, pois estas apresentam maiores restrições quanto à dispersão, o que a torna um fator importante na composição das comunidades (Hubbell, 1999; Chave, 2004).
Outros estudos mostram que a variação na similaridade de espécies também é associada a fatores ambientais, tais como clima, topografia, altitude e solos, sendo muito difícil separar os efeitos destes fatores mutuamente. Nesse caso, as comunidades são estruturadas pelo nicho (Tuomisto et al., 1995; Tuomisto & Poulsen, 1996; Ruokolainen & Tuomisto, 2002; Duivenvoorden et al., 2002; Ter Steege et al., 2002; Davidar et al., 2007), e a distribuição das espécies de plantas está então relacionada à distribuição espacial heterogênea dos diversos fatores ambientais mencionados (Pyke et al., 2001; Zuquim et al., 2007).
Duivenvoorden et al. (2002) afirmam que a heterogeneidade ambiental é um fator determinante na composição do conjunto de espécies em escala local, mas é necessário investigar-se até que ponto esse fator contribui para a manutenção da diversidade de plantas na escala de paisagem.
Na Amazônia Central, Costa et al. (2005) encontraram uma alta variação na composição de espécies de herbáceas em escala de 5 a 10 km (mesoescala), sendo que parte dessa variação foi relacionada à distância, consequência de limitações na dispersão das espécies.
Apesar da existência de diversos estudos demonstrar a diminuição da similaridade de espécies com o aumento da distância geográfica em grandes escalas (Ruokolainen & Tuomisto, 2002; Condit et al., 2002; Davidar et al., 2007), um estudo deveria ser feito no intuito de examinar se a distância geográfica influencia na variação da composição de espécies arbóreas em uma escala local (média de 0,2 a 10 km), pois diferentes padrões podem emergir em diferentes escalas. O conhecimento sobre a variação na composição das comunidades de espécies vegetais em diferentes escalas, da local à regional, é fundamental para o planejamento da conservação da biodiversidade em regiões tropicais (Pearman & Weber, 2007).
Este estudo tem como objetivo investigar se a Teoria da Neutralidade explica a variação na composição de espécies de árvores em uma floresta ombrófila na Amazônia oriental, em uma escala local.
MATERIAL E MÉTODOS
ÁREA DE ESTUDO
Este estudo foi realizado em um dos platôs da Floresta Nacional (FLONA) Saracá-Taquera, no distrito de Porto Trombetas, município de Oriximiná, Pará (Figura 1).
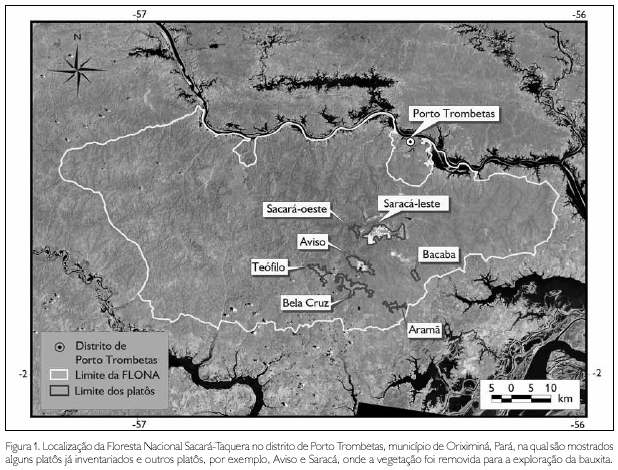
Nesses platôs ocorre a extração de bauxita pela Mineração Rio do Norte. A lavra do minério é feita a céu aberto, que é encontrado de 4 a 10 metros de profundidade, sob o solo da floresta ombrófila (Salomão et al., 2007).
Para a operação de desmatamento prévio, é necessário, por meio de um inventário botânico, avaliar a vegetação existente a fim de possibilitar o aproveitamento comercial da madeira e subsidiar as ações de recuperação florestal das áreas impactadas (Salomão et al., 2007).
O platô analisado neste estudo, denominado platô Bela Cruz, tem cerca de 1.500 hectares, sendo recoberto pela floresta ombrófila densa montana (Veloso et al., 1991), com topografia homogênea e altitude variando de 150 a 200 metros em relação ao nível do mar.
Predomina na área o latossolo amarelo distrófico, textura muito argilosa, e o latossolo amarelo distrófico, textura argilosa, sob floresta densa de relevo plano com bordos dissecados (Brasil, 1974).
A precipitação média anual na área varia de 2.500 mm a 3.000 mm, com dois períodos climáticos distintos: inverno, de janeiro a maio, quando ocorrem as maiores precipitações pluviométricas, e o verão, de junho a novembro, quando a estiagem é mais acentuada (Salomão et al., 2007).
As principais espécies de árvores do estrato emergente são: Dinizia excelsa Ducke (Mimosaceae), Bertholletia excelsa Bonpl. (Lecythidaceae), Tabebuia impetiginosa (Mart. ex DC.) Standl. (Bignoniaceae), Newtonia suaveolens (Miq.) Brenan (Mimosaceae) e Cedrelinga cateniformis (Ducke) Ducke (Salomão et al., 2007).
As principais espécies do dossel são Oenocarpus bacaba Mart. (Arecaceae), Geissospermum sericeum Benth. and Hook. f. ex Miers (Apocynaceae), Tetragastris panamensis (Engl.) Kuntze, Protium tenuifolium (Engl.) Engl. (Burseraceae) e Rinorea racemosa Kuntze (Violaceae) (Salomão et al., 2007).
Coleta de dados
No platô analisado foram distribuídas sistematicamente 179 parcelas de 250 m x 10 m, cobrindo a totalidade de sua superfície. Todas as parcelas foram georreferenciadas (Figura 2).

Dentro de cada parcela, todas as formas de vida vegetal (árvores, palmeiras e cipós) com Diâmetro à Altura do Peito (DAP a 1,30 m do solo) ≥ 10 cm foram medidas, marcadas e identificadas em nível taxonômico o mais específico possível.
Análise de dados
As curvas cumulativas de espécies e de estimadores de riqueza foram elaboradas através do programa Estimates 8 (Statiscal Estimation of species Richness and Shared Species form Samples) (Colwell & Coddington, 1996).
A matriz de similaridade de espécies entre as 179 parcelas foi gerada por meio do Índice de Sõrensen, cujo valor é baseado em termos da presença e ausência de espécies em pares de parcelas (Krebs, 1999), por meio do Programa Mata Nativa (Cientec, 2006). A matriz de distância geográfica entre as 179 parcelas foi calculada através do programa Arcview 3.3, usando a extensão "Distance Matrix of Point Features" (ESRI, 2002).
A correlação entre a similaridade de espécies e a distância linear entre as parcelas foi obtida pelo teste de Mantel, usando o índice de correlação de Pearson ao nível de significância de 5%, através do programa PC-ORD 4 (Zar, 2010).
O teste de Mantel é um procedimento estatístico de correlação entre duas matrizes (Mantel, 1967), frequentemente usado para correlacionar a variável 'distância' com outra variável qualquer (Manly, 1986). O método de Monte Carlo, com 1.000 permutações aleatórias, foi aplicado para avaliar a significância do teste de Mantel (Zar, 2010).
RESULTADOS
AMOSTRAGEM E RIQUEZA DE ESPÉCIES
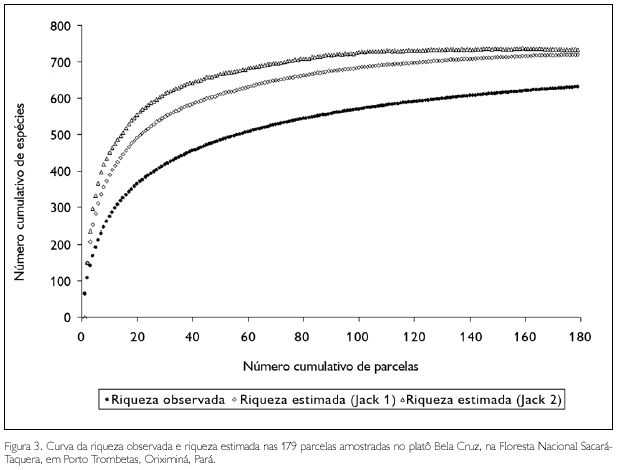
Nas 179 parcelas amostradas no platô, foram identificadas 631 espécies. As estimativas de riqueza de espécies, utilizando os estimadores de riqueza Jackknife 1 e 2, foram de 720 e 733 espécies, respectivamente, representado 88% e 86% da riqueza total observada, respectivamente, demonstrando que a amostragem realizada no platô foi satisfatória (Figura 3).
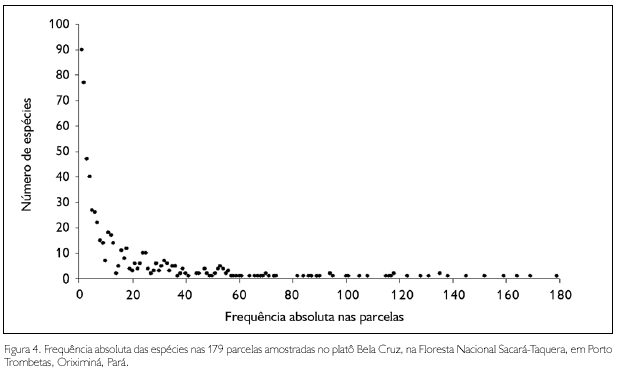
A maioria das espécies apresentou baixa frequência absoluta nas parcelas, pois entre as 631 espécies, 90 (14% do total) ocorreram em somente uma parcela, 77 (12% do total) ocorreram em duas parcelas e 442 espécies (70% do total) ocorreram em menos de 10% do total das parcelas (Figura 4).
SIMILARIDADE DE ESPÉCIES E DISTÂNCIA GEOGRÁFICA DAS PARCELAS
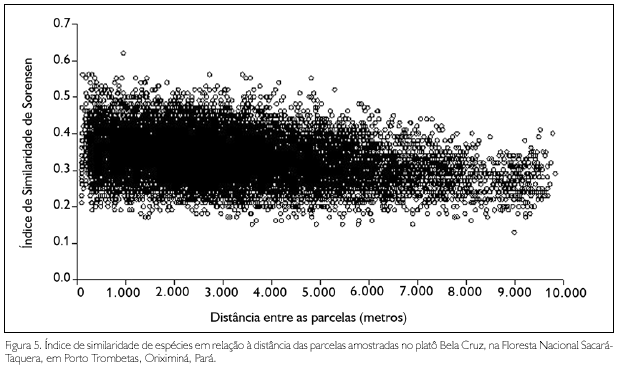
Houve uma correlação negativa significativa entre a similaridade de espécies e a distância geográfica entre as parcelas no platô (r = -0.44, t = 11,02; p = 0.0001, Figura 5).
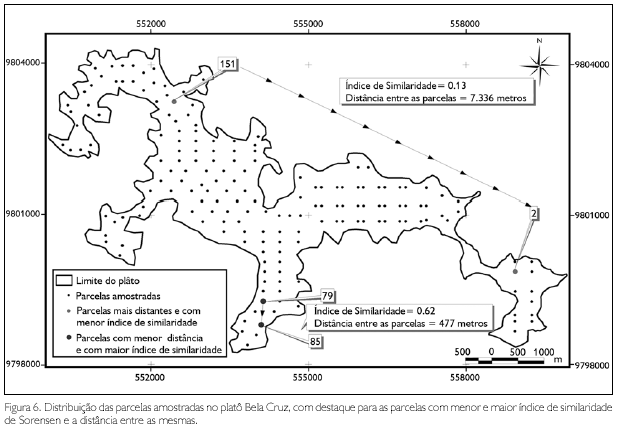
A distância entre as parcelas no platô variou de 200 metros a mais de 9 quilômetros. A menor similaridade de espécies (13%) ocorreu entre as parcelas 2 e 151, distantes 7,3 quilômetros entre si, enquanto a maior similaridade de espécies (62%) foi registrada entre as parcelas 79 e 85, distantes 477 metros entre si (Figura 6).
DISCUSSÃO
A correlação negativa entre a similaridade de espécies e a distância geográfica entre as parcelas no platô Bela Cruz ajusta-se ao estimado pela Teoria da Neutralidade (Hubbell, 2001), que prediz que a similaridade de espécies em uma comunidade diminui com o aumento da distância geográfica entre locais, independente de diferenças ambientais. Ou seja, como resultado de limitações na dispersão das espécies (Hubbel, 2006), quanto mais distantes as parcelas estiverem entre si, mais distintas elas serão em termos de sua composição florística.
Hubbell (2001) assume que a distribuição das espécies de plantas em uma comunidade é resultante de processos de imigração e extinção locais, onde estas espécies aparecem e desaparecem aleatoriamente. Com isso, espera-se que a variação na abundância de uma determinada espécie apresente forte correlação espacial devido às limitações de sua dispersão, ou seja, em comunidades tropicais há uma diminuição da similaridade de espécies vegetais em relação à distância geográfica entre as áreas amostradas.
Como resultado desse padrão, as comunidades de plantas na Amazônia são caracterizadas pelo grande número de espécies (Oliveira & Mori, 1999) e elevada proporção de espécies raras (Alves & Miranda, 2008).
Os mecanismos associados à limitação de dispersão indicam que altas concentrações de espécies raras em florestas tropicais estão associadas à propensão que estas espécies têm à extinção local, sendo substituídas mais lentamente na comunidade do que as espécies mais comuns (Hubbell, 2001; Maurer & Mcgill, 2005; Volkov et al,, 2005).
A densidade de algumas espécies tende a ser semelhante em locais próximos aos seus centros de dispersão, e sua densidade vai diminuindo à medida que a distância geográfica das áreas fonte aumenta. É evidente, no entanto, que quanto menor a taxa de dispersão das espécies, maior será a influência da distância geográfica em variações locais e regionais (Hubbell, 1999), porque há um decréscimo nas trocas de indivíduos entre locais geograficamente mais distantes.
Neste estudo, apesar da similaridade florística estar correlacionada negativamente com a distância geográfica (r = -0,44), demonstrando a sua influência na distribuição das espécies no platô, dentro de uma formação florestal aparentemente homogênea (como o platô Bela Cruz) pode existir microvariações das condições abióticas, que caracterizam habitats pontuais, por exemplo, relevo, e que não foram aqui testados. Por outro lado, vários estudos (Clark et al., 1999; Campbell & Peart, 2000; Richard et al., 2000; Svenning, 2001; Harms et al., 2004; Costa et al., 2005; Jones et al., 2006) demonstram que muitas espécies de plantas tropicais são distribuídas de acordo com variações de média escala das condições ambientais .
Chust et al. (2006) estudaram espécies de árvores em uma floresta tropical no Panamá e observaram que 22% da variação florística da área foi explicada pela distância geográfica entre os os locais de amostragem, 12% exclusivamente por variáveis ambientais e 16% pela correlação entre a distância geográfica dos locais de amostragem e as variáveis ambientais. Contudo, a maior parte da variação florística (49%) não foi explicada por nenhum destes fatores. Isto demonstra que há necessidade de mais estudos antes que compreendamos precisamente quais os eventos que determinam a variação da diversidade florística nestas florestas. Determinar a influência de diferentes processos na composição das comunidades de plantas tem importante aplicação na definição de planos de conservação mais eficientes em cada região (Pearman & Weber, 2007).
Infelizmente, as estratégias de conservação da biodiversidade em regiões tropicais são limitadas pelo ainda reduzido nível de conhecimento existente sobre a distribuição das espécies. Desta forma, há uma grande dificuldade em escolher as espécies para a definição de áreas prioritárias para a conservação da biodiversidade, pois o conhecimento atual sobre a riqueza, diversidade e endemismo de diversos grupos taxonômicos na Amazônia ainda é limitado.
CONCLUSÕES
No caso da Amazônia, propomos que as espécies consideradas raras devam ser também consideradas como espécies prioritárias para a conservação da biodiversidade, pois a floresta amazônica não é homogênea, mesmo em pequenas escalas (Tuomisto et al., 1995).
A diminuição da similaridade de espécies em relação à distância geográfica entre as parcelas inventariadas neste estudo é resultado da grande quantidade de espécies raras encontradas no platô Bela Cruz.
Apesar de diversos estudos terem demonstrado que a Amazônia é caracterizada pela alta riqueza e diversidade de espécies de plantas, poucos estudos têm levado em consideração a importância das espécies raras na conservação da biodiversidade na Amazônia.
O planejamento de estratégias de conservação na FLONA Saracá-Taquera deve considerar não somente a riqueza e diversidade locais de espécies, mas principalmente a conservação das espécies raras, pois, sem isto, corre-se o risco da eliminação da variação de espécies.
O padrão da similaridade de espécies no platô Bela Cruz mostra que a conservação da riqueza e diversidade de espécies da biota deve ser garantida pela conservação de áreas no platô. Por exemplo, na FLONA Saracá-Taquera, no platô Aviso, as porções leste e oeste foram deixados intactos. Esta estratégia pode ser repetida em outros platôs.
O planejamento de estratégias para conservação na FLONA Saracá-Taquera deve considerar a riqueza e diversidade locais de espécies, associada à variação desses parâmetros em uma escala regional, pois estratégias de conservação limitadas a escalas locais podem causar a eliminação da variação de espécies em escalas regionais.
AGRADECIMENTOS
À Mineração Rio do Norte (MRN), financiadora deste estudo; ao Instituto Chico Mendes de Biodiversidade (ICMBio), gestor da Floresta Nacional de Saracá-Taquera; às Dras. Veridiana V Scudeller, da Universidade Estadual do Amazonas (UEA), e Ana L. K. M. Albernaz, do Museu Paraense Emílio Goeldi (MPEG), pela ajuda na aplicação do uso de matrizes no Programa PC-ORD 4; e ao pesquisador Hanna Maoh, do Applied Research Institute-Jerusalem (ARIJ), pelo desenvolvimento e disponibilização da extensão "Distance Matrix of Point features", utilizada na geração da matriz de distâncias entre as parcelas deste estudo.
REFERÊNCIAS
ALVES, J. C. Z. O. & I. S. MIRANDA, 2008. Análise da estrutura de comunidades arbóreas de uma floresta amazônica de Terra Firme aplicada ao manejo florestal. Acta Amazonica 38(4): 657-666.
BRASIL, 1974. Departamento Nacional da Produção Mineral. Projeto RADAM. Folha SA 22 Belém. DNPM (Levantamento de Recursos Naturais, 5), Rio de Janeiro.
CAMPBELL, O. W. & D. R. PEART, 2000. Habitat association of trees and seedlings in a Bornean rain forest. Journal of Tropical Ecology 88: 464-478.
CARNEIRO, J. S. & D. M. VALERIANO, 2003. Padrão espacial da diversidade beta da Mata Atlântica – uma análise da distribuição da biodiversidade em um banco de dados geográficos. Anais do Simpósio Brasileiro de Sensoriamento Remoto 11: 629-636.
CHAVE, J., 2004. Neutral theory and community ecology. Ecology Letters 7: 241-253.
CHUST, G., J. CHAVE, R. CONDIT, S. AGUILAR, S. LAO & R. PÉREZ, 2006. Determinants and spatial modeling of tree beta – diversity in a tropical forest landscape in Panama. Journal of Vegetable Science 17: 83-92.
CLARK, D., M. W. PALMER & D. A. CLARK, 1999. Edaphic factors and the landscape-scale distributions of tropical rain forest trees. Ecology 80(8): 2662-2675.
COLWELL, R. K. & J. A. CODDINGTON, 1996. Estimating terrestrial biodiversity through extrapolation. In: D. L. HAWKSWORTH (Ed.): Biodiversity. Measurement and evaluation: 101-118. Chapman & Hall, London.
CONDIT, R., N. PITMAN, E. G. LEIGHT JR., J. CHAVE, J. TERBORGH, R. B. FOSTER, P. NÚÑEZ, S. AGUILAR, R. VALENCIA, G. VILLA, H. C. MULLER-LANDAU, E. LOSOS & S. P. HUBBELL, 2002. Beta-diversity in Tropical Forest Trees. Science 295(5555): 666-669.
CONSULTORIA E DESENVOLVIMENTO DE SISTEMAS (CIENTEC), 2006. Mata nativa 2: Manual do usuário: 1-295. Cientec, Viçosa.
COSTA, F. R. C., W. E. MAGNUSSON & R. C. LUIZAO, 2005. Mesoscale distribution patterns of Amazonian understorey herbs in relation to topography, soil and watersheds. Journal of Ecology 93(5): 863-878.
DAVIDAR, P., B. RAJAGOPAL, D. MOHANDASS, J-P. PUYRAVAUD, R. CONDIT, S. J. WRIGHT & E. G. LEIGH JR., 2007. The effect of climatic gradients, topographic variation and species traits on the beta diversity of rain forests trees. Global Ecology and Biogeography 16(4): 510-518.
DUIVENVOORDEN, J. F., J. C. SVENNING & S. J. WRIGHT, 2002. Beta Diversity in Tropical Forests. Science 295(5555): 636-637.
ENVIRONMENTAL SYSTEMS RESEARCH INSITUTE (ESRI), 2002. Arc View User’s Guide. Redlands, California.
HARMS, K. E., J. S. POWERS & R. A. MONTGOMERY, 2004. Variation in small sapling density, understory cover, and resource avaibility in four neotropical forests. Biotropica 36(1): 40-51.
HUBBELL, S. P., 1999. Light-Gap disturbances, recruitment limitation, and tree diversity in a neotropical forest. Science 283(5401): 554-557.
HUBBELL, S. P., 2001. The united neutral theory of biodiversity and biogeography. Princeton University Press, Princeton.
HUBBELL, S. P., 2006. Neutral theory and the evolution of ecological equivalence. Ecology 87: 1387-1398.
JONES, M. M., H. TUOMISTO, D. B. CLARK & P. OLIVAS, 2006. Effects of mesoscale environmental and dispersal limitation on floristic variation in rain forest ferns. Journal of Ecology 94: 181-195.
KNEITEL, J. M. & J. M. CHASE, 2004. Trade-offs in community ecology: linking spatial scales and species coexistence. Ecology Letters 7(1): 69-80.
KREBS, C. J., 1999. Ecological methodology: 2nd ed.: 1-620. Benjamin Cummings, Menlo Park, California.
MACARTHUR, R. H. & R. LEVINS, 1964. Competition, habitat selection and character displacement in a partchy environment. Proceedings of National Academy of Sciences 51(6): 1207-1210.
MANLY, B. F. J., 1986. Randomization and regression methods for testing for associations with geographical, environmental and biological distances between populations. Researches on Population Ecology 28(2): 201-218.
MANTEL, N., 1967. The detection of disease clustering and a generalized regression approach. Cancer Research 27: 209-220.
MAURER, B. A. & B. J. MCGILL, 2005. Neutral and non-neutral macroecology. Basic and Applied Ecology 5: 413-422.
OLIVEIRA, A. A. & S. A. MORI, 1999. A central Amazonian terra firme forest. I. High tree species richness on poor soils. Biodiversity and Conservation 8: 1219-1244.
PEARMAN, P. B. & D. WEBER, 2007. Common species determine richness patterns in biodiversity indicator taxa. Biological Conservation 138(1-2): 109-119.
PYKE, C. R., R. CONDIT, S. AGUILAR & S. LAO, 2001. Floristic composition across a climatic gradient in a Neotropical Lowland Forest. Journal of vegetation Science 12: 553-566.
RICHARD, M., T. BERNHARDT & G. BELL, 2000. Environmental heterogeneity and the spatial structure of fern species diversity in one hectare of old-growth forest. Ecography 23(2): 231-245.
RUOKOLAINEN, K. & H. TUOMISTO, 2002. Beta-Diversity in Tropical Trees. Science 297(5586): 1439.
SALOMÃO, R. P., N. A. ROSA & K. A. C. MORAIS, 2007. Dinâmica da regeneração natural de árvores em áreas mineradas na Amazônia. Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais 2(2): 85-139.
SCUDELLER, V. V., F. R. MARTINS & G. J. SHEPHERD, 2001. Distribution and abundance of arboreal species in the atlantic ombrophilous dense forest in Southeastern Brazil. Plant Ecology 152(2): 185-199.
SVENNING, J. C., 2001. On the role of microenvironmental heterogeneity in the ecology and diversification of neotropical rainforest palms (Arecaceae). The Botanical Review 67(1):1-53.
TILMAN, D., 2004. Niche tradeoffs, neutrality, and community structure: a stochastic theory of resource competition, invasion, and community assembly. Proceedings of the National Academy of Sciences of the United States of America 101(30): 10854-10861.
TUOMISTO, H., K. RUOKOLAINEN, R. KALLIOLA, A. LINNA, W. DANJOY & Z. RODRIGUEZ, 1995. Dissecting Amazonian Biodiversity. Science 269(5220): 63-66.
TUOMISTO, H. & A. D. POULSEN, 1996. Influence of edaphic specialization on pteridophyte distribution in neotropical rain forests. Journal of Biogeography 23(3): 283-293.
TER STEEGE, H., N. PITMAN, D. SABATIER, H. CASTELLANOS, P. V. D. HOUT, D. C. DALY, M. SILVEIRA, O. PHILLIPS, R. VASQUEZ, T. V. ANDEL, J. DUIVENVOORDEN, A. A. OLIVEIRA, R. EK, R. L. WAH, R. THOMAS, J. VAN ESSEN, C. BAIDER, P. MAAS, S. MORI, J. TERBORGH, P. N. VARGAS, H. MOGOLLÓN & W. MORAWETZ, 2002. A spatial model of tree α-diversity and tree density for the Amazon. Biodiversity and Conservation 12: 2255-2277.
VELOSO, H. P., A. L. R. RANGEL FILHO & J. C. A. LIMA, 1991. Classificação da vegetação brasileira adaptada a um sistema universal: 1-124. IBGE, Rio de Janeiro.
VOLKOV, I., J. R. BANAVAR, F. HE, S. P. HUBBELL & A. MARITAN, 2005. Density dependence explains tree species abundance and diversity in tropical forests. Nature 438: 658-651.
WRIGHT, J. F., M. T. FURSE, P. D. ARMITAGE & D. MOSS, 1993. New procedures for identifying running-water sites subjects to environmental stress and for evaluating sites for conservation, based on the macroinvertebrate fauna. Archiv fur Hydrobiologie 127(3): 319-326.
WRIGHT, S. J., 2002. Plant diversity in tropical forests: a review of mechanisms of species coexistence. Oecologia 130: 1-14.
ZAR, J. H., 2010. Biostatistical analysis: 5th ed.: 1-499. Prentice-Hall, Englewood Cliffs, New Jersey.
ZUQUIM, G., F. R. C. COSTA & J. PRADO, 2007. Fatores que determinam a distribuição de espécies de pteridófitas da Amazônia Central. Revista Brasileira de Biociências 5(2): 360-362.
 Autor para correspondência:
Autor para correspondência:
Leandro Valle Ferreira.
Museu Paraense Emílio Goeldi.
Coordenação de Botânica.
Avenida Perimetral, 1901.
Terra Firme. Belém, PA, Brasil.
CEP 66077-530
(lvferreira@museu-goeldi.br)
Recebido em 10/05/2010
Aprovado em 28/11/2011
Responsabilidade editorial: Toby Gardner
