










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkBoletim do Museu Paraense Emílio Goeldi Ciências Naturais
versão impressa ISSN 1981-8114
Bol. Mus. Para. Emilio Goeldi Cienc. Nat. v.7 n.1 Belém abr. 2012
Análise fitossociológica de floresta ombrófila densa e determinação de espécies-chave para recuperação de área degradada através da adequação do índice de valor de importância
Phytosociological analysis of tropical rain forest and determination of key-species to recovery of degraded area based through the importance value index
Rafael de Paiva SalomãoI; Antônio Cordeiro SantanaII; Silvio Brienza JúniorIII; Vitor Hugo Freitas GomesIV
IMuseu Paraense Emílio Goeldi/Ministério da Ciência Tecnologia e Inovação. Coordenação de Botânica. Belém, Pará, Brasil
IIUniversidade Federal Rural da Amazônia. Instituto Sócio-Ambiental e de Recursos Hídricos. Belém, Pará, Brasil
IIIEmbrapa Amazônia Oriental. Belém, Pará, Brasil
IVUniversidade Federal do Pará. Núcleo de Meio Ambiente. Belém, Pará, Brasil
RESUMO
Na Floresta Nacional Saracá-Taquera, estado do Pará, a lavra da bauxita é feita a céu aberto. As operações de lavra envolvem desde a supressão da floresta ombrófila densa até a restauração das áreas através de reflorestamentos heterogêneos, onde são empregadas por volta de 80 espécies. Objetivou-se, neste trabalho, apresentar um índice de fitossociologia horizontal (IFH), obtido pelas mesmas variáveis que fornecem o índice de valor de importância (IVI), através do ranqueamento das espécies amostradas nos inventários da floresta ombrófila densa em categorias ecológicas de prioridade alta, intermediária e baixa. Nos inventários fitossociológicos do platô Saracá, foram registrados 23.166 indivíduos, totalizando 796 espécies e 58 famílias. A adequação da análise fatorial foi determinada pelos testes de Bartlett, que avaliaram a significância geral da matriz de correlação, indicando que as correlações são significantes no nível de 1% de probabilidade, e de Kaiser-Meyer-Olkin (KMO), que indicou que as variáveis estão correlacionadas e que o modelo fatorial apresentou um nível bom de adequação aos dados. Os testes estatísticos validaram a amostra de dados para emprego da técnica de análise multivariada e, portanto, para a construção do índice de fitossociologia horizontal com variável dummy - IFH, que selecionou 65 espécies-chave para plantio em trabalhos de restauração florestal, contra apenas seis indicadas pelo índice IVI.
Palavras-chave: Seleção de espécies. Floresta tropical densa. Restauração florestal. Mineração de bauxita. Análise multivariada. Amazônia.
ABSTRACT
In the National Forest of Saracá-Taquera, Para State, the bauxite mining is made in open pits. The mining operations involve since the complete removal of the tropical rain forest to the land restoration by heterogeneous reforestation where are employed on average 80 species. The objective of this paper is to provide an index of horizontal phytosociology dummy (IHP), calculated through the same variables used to obtain the importance value index (IVI), by ranking tree species sampled in the inventories of the tropical rain forest in ecologic categories of high, medium and low priorities. The phytosociological inventories done in the Saracá plateau recorded 23,166 individuals, summing 796 species and 58 families. The adequacy of the factor analysis was determined by the tests of Bartlett, which evaluated the overall significance of the correlation matrix and shows that the correlations are significant to the 1% probability level, and Kaiser-Meyer-Olkin (KMO), which shows that the variables are correlated and that the factor model has a good fitness to the data set. The statistical tests validated the data set for use of multivariate analysis technique, and therefore for the construction of the IHP which selected 65 key species for use in forest restoration purposes, against only six indicated by the IVI.
Keywords: Species selection. Dense tropical forest. Forest restoration. Bauxite mining. Multivariate analysis. Amazonia.
INTRODUÇÃO
Em 1979, a Mineração Rio do Norte (MRN) iniciou as operações de lavra da bauxita na Floresta Nacional (FLONA) Saracá-Taquera/ICMBio, em Porto Trombetas, estado do Pará. A lavra do minério, que ocorre no subsolo da floresta ombrófila densa dos diversos platôs da área, é feita a céu aberto. As operações de lavra envolvem desde a supressão (corte raso) de toda a vegetação até a completa restauração florestal das áreas de lavra por meio dos reflorestamentos heterogêneos, onde são empregadas por volta de 80 espécies arbóreas, e a indução da regeneração natural, via adição e espalhamento de solo superficial (top soil, 'terra preta') nas áreas anuais de plantio. Antes da operação de supressão, a empresa tem que avaliar quali e quantitativamente a vegetação florestal, por meio de inventários florestais e fitossociológicos, para possibilitar o aproveitamento de toda a madeira comercial e também subsidiar as ações de restauração florestal das áreas desmatadas para a extração de bauxita.
A ação antrópica é necessária para a recuperação desses ecossistemas degradados pela atividade minerária, pois já não dispõem de mecanismos naturais de regeneração, tais como banco de sementes, banco de plântulas, chuva de sementes e rebrota, apresentando, portanto, baixa resiliência, isto é, seu retorno ao estado anterior pode ou não ocorrer ou ser extremamente lento (Carpanezzi et al., 1990). Nos processos de recuperação destes ecossistemas, podem ser usadas duas técnicas distintas: restauração e reabilitação.
Restauração, de acordo com Rodrigues et al. (2009), é o retorno de uma condição perturbada ou totalmente alterada a um estado anterior que existia naturalmente. Moscatelli et al. (1993) afirmaram que a restauração refere-se ao retorno a um estado pré-existente sem que, necessariamente, o sistema retorne às suas características originais. Segundo Viana (1990), a restauração refere-se ao conjunto de tratamentos que visam recuperar a forma original do ecossistema, ou seja, a sua estrutura, dinâmica e interações biológicas, sendo uma técnica recomendada para ecossistemas raros e ameaçados, nos quais os processos de recuperação demandam maior tempo e resultam em custos elevados (caso das áreas de mineração). A restauração ecológica de áreas degradadas envolve o atendimento a pelo menos nove atributos listados pela Society for Ecologial Restoration (SER, 2004), entre os quais se destaca que o ecossistema restaurado deve conter um conjunto característico de espécies-chave ocorrentes no ecossistema de referência, fornecendo uma estrutura apropriada de comunidade, e também que seja constituído pelo maior número possível de espécies nativas.
Os platôs da FLONA Saracá-Taquera/ICMBIO, em especial, o platô Saracá, objeto deste estudo, podem ser considerados como o extremo da degradação antrópica, pois todas as propriedades físicas, químicas, biológicas e ecológicas do ecossistema original foram intensamente alteradas ou destruídas após a lavra da bauxita, acarretando uma profunda instabilidade de todo o ecossistema artificial criado (Salomão et al., 2007).
Na base da definição dos métodos adequados para a restauração florestal, uma questão é fundamental para o sucesso do objetivo, independentemente do método selecionado: quais espécies plantar de modo a recobrir o solo e restaurar os processos ecológicos em menos tempo, com menores perdas e ao menor custo. A seleção das espécies adequadas é de fundamental importância para o sucesso da restauração. A grande maioria, se não a totalidade, dos Planos de Recuperação de Áreas Degradadas (PRAD) das empresas minerárias indica que se use as espécies de maior índice de valor de importância (IVI), descrito por Curtis & McIntosh (1951). No entanto, não existe nenhum estudo para a Amazônia que seja fundamentado em uma análise amparada por modelos estatísticos robustos e indique quais espécies plantar.
Um importante instrumento da ecologia de comunidade é a análise multivariada, que trata todas as variáveis simultaneamente, sumariando os dados e revelando a sua estrutura com a menor perda de informações possível (Gauch, 1982; Pielou, 1984). Diferentemente da estatística clássica, que possibilita o teste de hipótese, as análises multivariadas se prestam mais a investigar os dados e gerar hipóteses (Gauch, 1982; Oliveira-Filho & Ratter, 1995). Só recentemente as análises multivariadas tornaram-se instrumentos acessíveis, apesar de há muito conhecidas (Goodall, 1954). Muitos estudos em comunidades vegetais utilizam como base esse tipo de análise, seja para agrupar amostras, classificar tipos de formações, relacionar variáveis ambientais a diferenças na comunidade, ou mesmo para definir prioridades para a conservação (Toledo et al., 2009; Santos et al., 2007; Johnson & Wichern, 2007; Stella, 2002; Silva & Shepherd, 1986).
A análise multivariada foi empregada por Toledo et al. (2009) para caracterização fitossociológica em vegetação de cerrado no norte de Minas Gerais. Santos et al. (2007) analisaram a riqueza e a similaridade florística de fragmentos florestais no norte de Minas Gerais baseando-se no dendrograma e na distância euclidiana entre as parcelas, calculada a partir da matriz de correlação entre as variáveis selecionadas. Stella (2002) testou uma metodologia de seleção de espécies arbóreas tropicais prioritárias para a conservação, a partir de um total de 214 espécies lenhosas conhecidas, sendo cinco delas selecionadas; concluiu informando que a análise do sistema dos algoritmos, bem como a da seleção e uso dos critérios, revelou pontos fortes e fracos da metodologia. Silva e Shepherd (1986) estabeleceram relações entre diferentes amostras de mata atlântica e floresta amazônica, utilizando a análise de agrupamento no nível de gêneros.
Para estudos fitossociológicos, o estabelecimento de parcelas na área que se deseja estudar é o procedimento mais comumente empregado. Usa-se a média das contagens e mensurações feitas para cada espécie em cada parcela, e supõe-se que as médias assim obtidas possam ser generalizadas para toda a fitocenose. Esses métodos permitem avaliar quantitativamente a variabilidade dos parâmetros estimados e também podem fornecer, simultaneamente, informações sobre o padrão espacial de distribuição de indivíduos em cada população. O trabalho de Cain et al. (1956) marcou o início da utilização, na Amazônia, de parâmetros fitossociológicos como densidade, frequência e valor de importância (Curtis & Mcintosh, 1951) e da metodologia de caracterização da vegetação de Raunkiaer (1934), utilizando formas de vida e tamanho de folha.
Prance et al. (1976) produziram o primeiro, e por muito tempo o único, estudo quantitativo da floresta de terra firme da região, utilizando coletas para identificações no nível de espécie. Rankin-de-Merona et al. (1992) publicaram os dados preliminares do levantamento de 70 ha de mata de terra firme próximo a Manaus, dentro das reservas do Projeto Dinâmica Biológica de Fragmentos Florestais (PDBFF), onde foram computadas 698 morfoespécies, com as famílias Lecythidaceae, Leguminaceae, Sapotaceae e Burseraceae. Essas famílias foram as mais abundantes em número de indivíduos e também as mais ricas em espécies. Valencia et al. (1994) amostraram, no Equador, 1 ha de floresta ombrófila com a maior alfa-diversidade de árvores do mundo. Foram computadas 307 espécies com DAP ≥ 10 cm, sendo que Jessenia bataua (Arecaceae) e Eschweilera aff. coriacea (Lecythidaceae) foram as espécies com maior densidade, e Leguminosae, Lauraceae, Sapotaceae, Moraceae e Burseraceae, as famílias mais diversas. Assim, os inventários na Amazônia têm demonstrado que as matas de terra firme apresentam alta diversidade, grande porcentagem de espécies com baixa densidade e baixa similaridade florística entre parcelas próximas. Esses padrões foram estabelecidos muito cedo na história dos inventários florestais quantitativos nessa região (Davis & Richards, 1934; Black et al., 1950; Pires et al., 1953; Cain et al., 1956), pouco tendo sido acrescentado desde então quanto a padrões estruturais e florísticos.
É muito difícil destacar algumas dezenas de espécies para a restauração, entre as centenas que ocorrem, de um modo geral, nos platôs da FLONA Saracá-Taquera/ICMBio, que devem apresentar uma grande importância na estrutura dessas florestas. O matá-matá (Eschweilera coriacea, Lecythidaceae) é a única espécie que aparece com grande densidade em levantamentos por toda a Amazônia (Valencia et al., 1994; Rollet, 1993; Silva et al., 1992; Prance et al., 1976; SUDAM, 1973; Cain et al., 1956; Pires et al., 1953). Contudo, alguns padrões bem estabelecidos emergem das análises no nível de família, na quais, com raras exceções, Leguminosae (sensu lato), Lecythidaceae, Sapotaceae, Burseraceae, Chrysobalanaceae, Moraceae e Lauraceae estão entre as famílias com maior densidade e diversidades no dossel de matas primárias.
A Secretaria de Meio Ambiente (SMA) do estado de São Paulo publicou uma legislação inédita para orientar a recuperação de áreas degradadas: a Resolução SMA 47, de 21/11/2003 (SMA, 2003), preconiza, entre outras normas, que nos trabalhos de restauração florestal, em áreas originalmente cobertas por florestas, deverá ser usado um número mínimo de 80 espécies. No estado do Pará, o Decreto no. 31.594, de 27/01/2010 (IDEFLOR, 2010), dispõe no artigo 7o, inciso III, que o plantio de mudas para fins de recomposição da Reserva Legal (80% da área total do imóvel) deverá utilizar o maior número possível de espécies nativas de ocorrência regional.
Considerando que as empresas minerárias têm que restaurar as áreas florestais cuja cobertura vegetal foi suprimida, objetivou-se, neste estudo, analisar a estrutura fitossociológica horizontal por meio da adequação de um índice fitossociológico com variável dummy (IFH), obtido por técnicas de análise multivariada, envolvendo as mesmas variáveis do índice de valor de importância (IVI) proposto por Curtis & McIntosh (1951). Posteriormente, foi feita a comparação dos resultados obtidos pelos dois métodos através da hierarquização das espécies amostradas na floresta ombrófila densa em três categorias ecológicas previamente estabelecidas (prioridade alta, intermediária e baixa). Após o ranqueamento, foi feita a seleção das espécies mais adequadas (espécies-chave) para o plantio nas áreas anuais de restauração florestal (média de 400 ha/ano), onde devem ser usadas um mínimo de 80 espécies.
MATERIAIS E MÉTODOS
ÁREA DE ESTUDO
Os inventários florestais foram realizados, no período de 2002 a 2006, em uma área total de 1.290,5 ha, no platô Saracá, com altitude média de 180 m, localizado na Floresta Nacional Saracá-Taquera/ICMBio, no distrito de Porto Trombetas (1o 21' S - 56o 22' W), município de Oriximiná, estado do Pará, Brasil.
De acordo com o Instituto Brasileiro de Geografia e Estatística (IBGE, 1992), a área de estudo acha-se inserida na região da Floresta Ombrófila Densa, sub-região dos Baixos Platôs da Amazônia, domínio da floresta densa das baixas altitudes, cuja fisionomia refere-se à floresta localizada principalmente nos platôs Terciários e terraços antigos e recentes, apresentando-se em dois estratos distintos: um emergente e outro uniforme. As principais espécies que caracterizam o estrato emergente são Dinizia excelsa (angelim-pedra), Bertholletia excelsa (castanheira) e Cedrelinga cateniformis (cedrorana). O estrato uniforme é caracterizado por Manilkara spp. (maçarandubas), Protium spp. (breus) e Pouteria spp. (abius).
PROCESSO DE AMOSTRAGEM
O processo amostral utilizado nos inventários florestais foi baseado na amostragem sistemática, também conhecida como seleção mecânica, onde as unidades amostrais são selecionadas segundo um rígido e pré-determinado esquema que procura cobrir toda a população (Campos & Leite, 2002).
O princípio básico do método consiste em selecionar as unidades amostrais (ou parcelas) seguindo um intervalo k, definido antecipadamente. A seleção da primeira parcela automaticamente determina a posição das demais. Assim sendo, o elemento chance pode ser introduzido somente no ponto de partida (Soares et al., 2006). Por outro lado, a direção a seguir pela linha imaginária (eixo) das parcelas muitas vezes é definida propositalmente, senão os resultados poderão ser influenciados, por exemplo, pela topografia. Procura-se, geralmente, o sentido da linha de declividade.
Atualmente, ainda perduram divergências quanto à precisão das estimativas provenientes da amostragem sistemática, sendo defendida a ideia de que somente as técnicas da seleção aleatória dos parâmetros da população oferecem estimativas isentas de tendência (Sanquetta et al., 2009). Não obstante, no presente estudo, utilizou-se uma concepção de amostragem sistemática com múltiplos inícios aleatórios, convergindo, assim, para um procedimento casualizado, tal como o fizeram Queiroz & Barros (1998). A amostragem sistemática, em muitos casos, produz estimativas confiáveis, além de ser bem menos onerosa devido ao modo como é esquematizada a localização das parcelas. A fixação das unidades amostrais é mais fácil e mais rápida, assim como a organização, a supervisão e a checagem tornam-se mais simples de serem executadas.
A variância da média, sob o ponto de vista teórico da amostragem sistemática, de acordo com Queiroz & Barros (1998), somente poderá ser obtida uma vez que se disponha dos valores médios de todas as amostras sistemáticas possíveis, ou no caso de serem conhecidos os valores verdadeiros da variância entre unidades dentro das amostras sistemáticas, o que corresponde a ter que enumerar ou levantar totalmente a população, o que não é factível em termos de inventário florestal. Os autores sugerem que, em face da impossibilidade de se obter a variância da média de uma única amostra sistemática, pode-se, relativamente aos levantamentos florestais, analisá-la como inteiramente casualizada, uma vez que as árvores estão distribuídas aleatoriamente por algum critério objetivo, ocorrendo na floresta segundo leis naturais.
A vantagem mais importante da amostra sistemática, quando aplicada em florestas primárias, é que as unidades amostrais se distribuem mais uniformemente na população, originando uma maior representatividade, tornando-se eficiente para detectar tendência ou concentração de certas características, sendo também mais precisa na observação das variáveis tipológicas (Queiroz & Barros, 1998; Campos & Leite, 2002; Soares et al., 2006; Sanquetta et al., 2009).
A unidade amostral empregada tinha a forma retangular, com 10 m de largura por 250 m de comprimento (0,25 ha), onde foram registradas, identificadas e mensuradas todas as árvores, cipós e palmeiras com DAP ≥ 10 cm. Foram empregadas 195 parcelas na amostragem, resultando em uma intensidade amostral de 3,78%.
O material fértil coletado está depositado no Herbário do Museu Paraense Emílio Goeldi (MPEG), com sigla internacional MG. A identificação do material botânico foi feita por comparação pelos parabotânicos do MPEG, sendo que o material fértil foi encaminhado aos especialistas. O sistema de classificação adotado foi o Angiosperm Phylogeny Group (APG iii).
ESTRUTURA HORIZONTAL
A estrutura horizontal da floresta foi caracterizada por meio das variáveis fitossociológicas incluídas na análise fatorial, como abundância, frequência e dominância (absolutas e relativas) das espécies registradas nas parcelas da amostra. Estas variáveis foram utilizadas na definição de um índice de fitossociologia horizontal (IFH) para ser comparado com o índice de valor de importância ecológica da espécie (IVI), proposto por Curtis & Mcintosh (1951). Alternativamente, incluiu-se uma variável qualitativa (variável dummy - VD) para captar a influência das espécies em que pelo menos 50% dos valores das variáveis estudadas constavam na amostra.
O IVI é definido de acordo com a equação IVI = ABR + FRR +DOR em que: (i) ABR = (número de indivíduos da espécie / número total de indivíduos x 100); (ii) FRR = (número de parcelas em que ocorre a espécie / soma do número total de parcelas de todas as espécies) x 100; (iii) DOR = (área seccional total dos indivíduos da espécie / área seccional total de todos os indivíduos das espécies) x 100.
Para a análise da riqueza e da diversidade arbórea, foram utilizados os índices de Shannon & Wiener (1949), de equabilidade (Pielou, 1975) e o índice de Simpson (Simpson, 1949).
MODELO ANALÍTICO
O modelo de análise fatorial, conforme Santana (2005, 2007, 2008), foi utilizado para definir as dimensões latentes comuns, ou fatores comuns, a partir da estrutura de inter-relações apresentadas entre as variáveis quantitativas e qualitativas da amostra de dados utilizada no trabalho.
A expressão matemática que especifica o modelo geral de análise fatorial, conforme apresentado em Dillon & Goldstein (1984), é dada por:
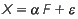
em que: (i) X = é o vetor transposto p-dimensional das variáveis observáveis, dado por X = (x1' x2' Xp), (ii) F é o vetor transposto q-dimensional das variáveis latentes ou fatores comuns, dado por F = (f1' f2' ..., fq), assumindo q < p; (iii) 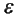 = é o vetor transposto p-dimensional das variáveis aleatórias ou fatores únicos, dado por
= é o vetor transposto p-dimensional das variáveis aleatórias ou fatores únicos, dado por 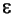 = (e1' e2' ..., ep); e (iv) α = é a matriz (p, q) dos parâmetros ou cargas fatoriais a serem estimados.
= (e1' e2' ..., ep); e (iv) α = é a matriz (p, q) dos parâmetros ou cargas fatoriais a serem estimados.
As cargas fatoriais geradas a partir da solução inicial do modelo de análise fatorial podem não representar um padrão significativo, caso não se consiga uma ortogonalidade perfeita entre elas. Para corrigir esta questão, conforme Dillon & Goldstein (1984) e Johnson & Wichern (1992), adotou-se o método varimax de rotação ortogonal dos fatores pela simplicidade e pelos bons resultados que apresenta (Hair et al., 2005; Santana, 2005).
A técnica de raiz unitária foi utilizada para a extração dos fatores (Hair et al., 2005; Mingoti, 2005; Santana, 2007, 2008). Sendo assim, as variáveis que contribuíram com um autovalor igual ou superior a 1 foram escolhidas, uma vez que somente estas são consideradas significantes para representar o fenômeno estudado.
Para a definição do índice fitossociológico horizontal com variável dummy (IFH), procedeu-se conforme Santana (2007), a partir da estimação dos escores fatoriais. Assim, para cada fator extraído fj' oi-ésimo escore fatorial extraído é definido por Fij' expresso conforme Dillon e Goldstein (1984):
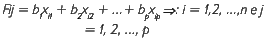
onde bi são os coeficientes de regressão estimados para os n escores fatoriais comuns e xij são as n observações das p variáveis observáveis, padronizadas pelo método Z-score para apresentar uma distribuição com média 0 e desvio igual a 1.
Para a construção do IFH, o escore fatorial (Fi) foi padronizado a fim de se obter valores positivos dos escores originais para variar entre 0 e 1 (FPi) e permitir a hierarquização das espécies. A fórmula matemática é a seguinte:
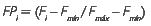
onde Fmax e Fmín são os valores máximo e mínimo observados para os escores fatoriais associados às variáveis (abundância, frequência e dominância) das espécies.
O IFH foi definido com base em Santana (2007):
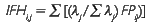
onde λ é a variância explicada por cada fator e ∑λ é a soma total da variância explicada pelo conjunto de fatores comuns extraídos.
Para facilitar a interpretação dos resultados, foram estabelecidos três intervalos de valores do IFH e do IVI, obtidos pela diferença entre o maior e o menor valores calculados, dividida por 3, aqui denominada amplitude de ranqueamento (AR).
Espécies de alta prioridade foram aquelas com índices situados no intervalo entre o maior valor do IFH calculado, subtraído da amplitude de ranqueamento, enquanto as espécies com prioridade baixa foram aquelas cujos índices calculados encontravam-se no intervalo entre o menor valor do IFH mais a amplitude de ranqueamento. As espécies cujos índices calculados situaram-se entre IFH + AR e IFH - AR foram classificadas como de prioridade intermediária; analogamente, tal procedimento foi estendido para os cálculos do IFH e IVI.
Na análise fatorial, as variáveis explicativas Xi (i = 1, 2, n) são quantitativas e qualitativas. A variável de natureza qualitativa incluída na análise teve a missão de identificar a importância de algumas espécies na amostra. Para isso, atribuiu-se valor 1 para indicar a presença da característica de interesse e valor 0 para revelar a ausência do fenômeno.
Os fenômenos analisados, cuja variável dummy assumiu valor igual a 1, referem-se àquelas espécies domi n an tes cu jos m ai ores valores acu mu lados e individuais da abundância, frequência e dominância absolutas responderam por mais de 50% do valor total de cada uma dessas variáveis. Por exemplo: a abundância absoluta de todas as espécies foi de x indivíduos; após a hierarquização decrescente das espécies, do maior para o menor valor desta variável, procedeu-se à soma do número de indivíduos de cada espécie até se atingir 50% do total de x e as espécies aí incluídas receberam valor 1 na variável dummy, enquanto as demais receberam o valor zero. Raciocínio análogo foi feito para a frequência e a dominância. Normalmente, essas variáveis constituem um número pequeno em relação ao tamanho da amostra.
Para aferir a adequação do método à amostra de dados, foram aplicados os testes de Kaiser-Meyer-Olkin (KMO), que compara as correlações entre as variáveis observáveis (Hair et al., 2005) e de esfericidade de Bartlett, que testa a hipótese nula de que a matriz de correlação é uma matriz identidade (Dillon & Goldstein, 1984; Gama et al., 2007). Os resultados foram obtidos por meio do software SPSS Statistics 17.0™.
RESULTADOS E DISCUSSÃO
FLORÍSTICA E DIVERSIDADE
Foram registrados nos inventários fitossociológicos do platô Saracá, nas 195 parcelas (0,25 ha cada, totalizando 48,75 ha de vegetação amostrada), 23.166 indivíduos (DAP ≥ 10 cm), totalizando 796 espécies, distribuídas em 58 famílias (Apêndice).
Os índices de diversidade calculados (Shannon e Wiener - H' = 5,438 e de Simpson, 1-D = 0,987), de dominância de Simpson (D = 0,013) e equabilidade (J = 0,812) entre as parcelas analisadas, mostram que a comunidade é altamente diversa. O índice de diversidade de espécies de Shannon e Wiener, de acordo com Margalef (1972), normalmente apresenta valores entre 1,5 a 3,5, raramente ultrapassando 4,5 para logaritmo neperiano. Todavia, Oliveira e Amaral (2004) registraram, em floresta de vertente na Amazônia Central, o valor de 5,01 para esse índice, considerado muito elevado dentro do contexto de levantamentos semelhantes na região. Salomão et al. (2007) apresentam resultados de mais de duas dezenas de registros para a Amazônia cujos índices variaram de 4,90 a 2,63; ressaltando que na floresta ombrófila densa do médio rio Xingu foram registrados valores de 4,78 em Senador José Porfírio (Pará), 4,64 em Anapu (Pará), e 4,40 em Altamira (Pará).
O índice de diversidade de Simpson (1-D) correspondeu a aproximadamente 1 (0,987). O índice de dominânica de Simpson (D) indicou que não há dominância de espécies nas parcelas inventariadas, ficando em 1,1% a probabilidade de se amostrar ao acaso dois indivíduos que pertençam à mesma espécie. A equabilidade é alta, indicando que os valores de diversidade de Shannon e Wiener são distantes da média esperada para o número de espécies amostradas.
A maior riqueza de espécies foi apresentada por Sapotaceae (97 espécies), Mimosaceae (56), Chrysobalanaceae (46), Lauraceae (45), Fabaceae (42), Annonaceae (39), Myrtaceae (37), Moraceae (35), Lecythidaceae (33), Burseraceae e Caesalpiniaceae (31 espécies cada); em toda a amostragem, oito famílias (ou 13,8% do total) ocorreram com apenas uma espécie. Considerando-se Leguminosae (sensu lato), esta seria a família com maior riqueza: 129 espécies.
Sapotaceae foi a família q ue apresentou o maior valor de abundância nos 48,75 ha da amostra (3.578 indivíduos ou 73,4 ind. ha-1), seguindo-se Violaceae (3.113 ou 63,9 ind. ha-1), Lecythidaceae (2.078 ou 42,6 ind. ha-1), Annonaceae (1.142 ou 23,4 ind. ha-1), Mimosaceae (1.089 ou 22,3 ind. ha-1), Euphorbiaceae (1.015 ou 20,8 ind. ha-1), Burseraceae (969 ou 19,9 ind. ha-1) e Chrysobalanaceae (834 ou 17,1 ind. ha-1); apenas estas oito famílias responderam por 59,6% (ou 13.818 espécimes) do total de indivíduos da amostra. Considerando-se Leguminosae (sensu lato), esta seria a família mais abundante, com 4.832 indivíduos ou 99,1 ind. ha-1.
Os gêneros com maior número de espécies foram Pouteria (Sapotaceae), Protium (Burseracae) e Eschweilera (Lecythidaceae), com, respectivamente, 2.621, 1.797 e 802 árvores; a maior abundância foi observada para Rinorea (Vilaceae), com 3.042 árvores na amostra.
As espécies de maior abundância na amostra foram Rinorea riana (1.741 indivíduos em 48,75 ha ou 35,7 ind. ha-1), Rinorea racemosa (942 ou 19,3 ind. ha-1), Eschweilera amazonica (656 ou 13,5 ind. ha-1), Eschweilera coriacea (613 ou 12,6 ind. ha-1), Geissospermum sericeum (523 ou 10,7 ind. ha-1), Oenocarpusbacaba (473 ou 9,7 ind. ha-1), Eschweilera grandiflora (376 ou 7,7 ind. ha-1), Rinorea guianensis (348 ou 7,1 ind. ha-1) e Hevea guianensis (286 ou 5,9 ind. ha-1); apenas estas nove espécies (ou 1,1% do total) ocorreram com 5.958 do total de indivíduos (ou 25,7% do total); no outro extremo, 137 espécies (ou 16,9% do total) ocorreram com apenas um indivíduo.
Nenhuma espécie ocorreu em todas as 195 unidades amostrais, consequentemente nenhuma apresentou 100% de frequência. Eschweilera coriacea, Geissospermum sericeum, Eschweilera amazonica, Rinorea racemosa, Oenocarpus bacaba, Eschweilera grandiflora e Rinorea riana ocorreram em 180 (92,3% do total), 178 (91,3%), 170 (87,2%), 168 (86,2%), 161 (82,6%), 152 (77,9%) e 151 (77,4%) parcelas, respectivamente; no outro extremo, 154 espécies (ou 18,9% do total) ocorreram em apenas uma parcela da amostra.
ÍNDICES FITOSSOCIOLÓGICOS
Os índices ecológicos, biológicos e fitossociológicos permitem uma melhor compreensão dos ecossistemas, sobretudo os tropicais, que apresentam uma grande diversidade biológica, classificando e agrupando as comunidades vegetais e as populações em categorias semelhantes, tornando mais simples a percepção e a integração das variáveis envolvidas nas análises e refletidas nos respectivos índices.
Índice de Fitossociologia Horizontal com Variável Dummy (IFH)
A variável dummy (VD), com valor igual a 1, foi atribuída àquelas espécies cuja abundância, frequência e dominância, classificadas separadamente do maior para o menor valor, apresentaram valor cumulativo da respectiva variável igual ou imediatamente superior a 50%, atingindo desta forma, metade do valor total da variável, ou seja, para a abundância, frequência e dominância foram registradas, respectivamente, 51, 16 e 48 espécies que compreendem mais de 50% do total da variável em questão.
A adequação da análise fatorial foi determinada pelos testes Bartlett e KMO. O teste de Bartlett avaliou a significância geral da matriz de correlação e apresentou um valor de 3.579,473, indicando que as correlações, em geral, são significantes ao nível de 1% de probabilidade, ou seja, a matriz de correlação não é diagonal. O teste KMO, da ordem de 0,829, indicou que as variáveis estão correlacionadas e o modelo fatorial apresentou um nível bom de adequação aos dados. Estes resultados respaldam o emprego da análise fatorial para a extração de fatores e a estimação dos escores fatoriais fitossociológicos.
Os resultados da solução inicial foram rotacionados para os três fatores pré-determinados, visando confirmar a adequação dos resultados obtidos com relação à significância das cargas fatoriais estimadas. Aplicando-se o critério da raiz latente, três componentes foram mantidas ou extraídas (Dillon & Goldstein, 1984). Observou-se que os três fatores explicaram 87,012% da variância total da nuvem de dados (Tabela 1), o que é amplamente explicado pelo critério da porcentagem da variância. Na referida tabela, a penúltima linha contém o resultado para a variância total explicada por cada uma das componentes, e na última linha estão os porcentuais da variância explicada pelas respectivas componentes: a componente 1 explicou 30,058% da variância total, a componente 2 explicou 28,837% e a componente 3 explicou 28,117%.
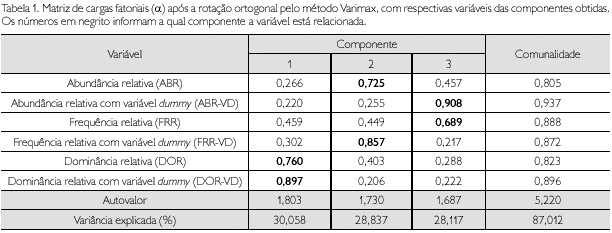
A estatística, detalhando o grau em que cada variável é explicada pelas três componentes, é denominada de Comunalidade, cuja soma do quadrado de cada fator está representado na respectiva coluna (Comunalidade) da Tabela 1; o número 5,22 representa o total das cargas fatoriais ao quadrado e indica a parcela total de variância extraída pela solução fatorial rotacionada, igual a 87,012%.
O índice IFH para esta solução mostrou que 87,012% da variância total são representados pela informação contida na matriz fatorial da solução em termos dos três fatores. O índice é considerado muito bom, e as variáveis estão, como esperado, estreitamente relacionadas umas com as outras.
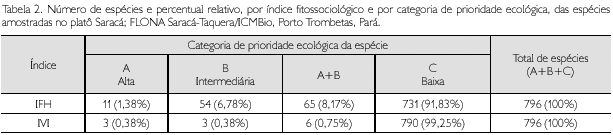
O IFH calculado variou de 0,357% a 0,107%, consequentemente, a amplitude de 0,250% dividida por 3 definiu a amplitude de ranqueamento (AR) de 0,083%, gerando os três intervalos das categorias pré-estabelecidas de prioridade das espécies: alta (0,357% a 0,274%), intermediária (0,273% a 0,190%) e baixa (0,189% a 0,107%). Foram registradas 11 espécies com prioridade alta, 54 como de categoria intermediária e as demais 731 como espécies de baixa prioridade (Tabela 2).
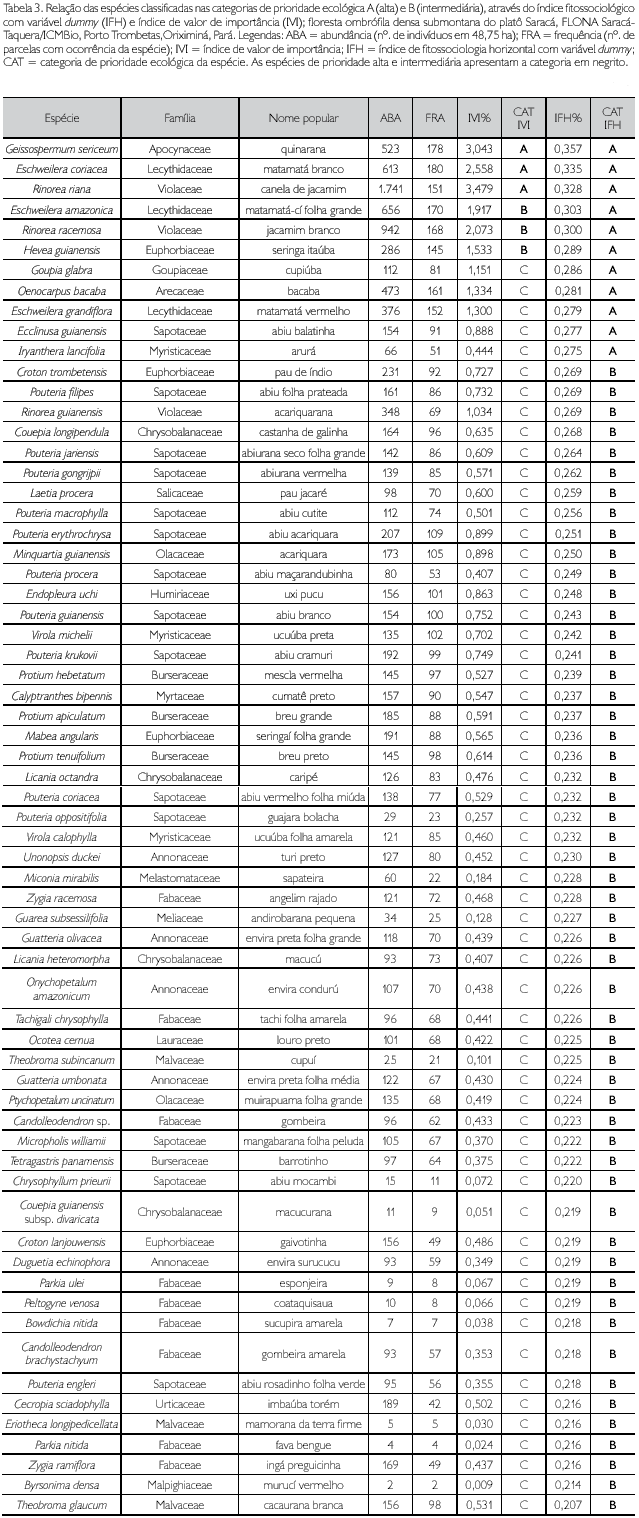
A relação de todas as espécies classificadas como de alta e intermediária prioridade é apresentada na Tabela 3.
Índice de Valor de Importância (IVI)
O IVI calculado para cada espécie variou de 3,479% a 0,349%, produzindo uma amplitude de 3,13% que, dividida por 3, resultou numa amplitude de ranqueamento (AR) igual a 1,043%, gerando as respectivas categorias pré-estabelecidas de prioridade ecológica das espécies: alta (de 3,479% a 2,436%), intermediária (de 2,435% a 1,393%) e baixa (de 1,392% a 0,349%). Foram registradas apenas três espécies nas categorias de prioridade alta e intermediária; a quase totalidade das demais 790 espécies (ou 99,25% do total) foi classificada como de prioridade baixa (Tabela 2). A relação nominal de todas as espécies classificadas como de alta e intermediária prioridades e de outras 59 de baixa prioridade é apresentada na Tabela 3.
Em termos de biodiversidade e de fatores socioeconômicos, e devido à grande riqueza e diversidade de espécies vegetais presentes nas florestas tropicais, a interpretação dos dados pode tornar-se um grande problema, e ao mesmo tempo ser uma grande vantagem. Tome-se como exemplo este trabalho, onde foram registradas 796 espécies, distribuídas em 58 famílias, totalizando 23.166 indivíduos (média de 475,2 árvores ha-1) presentes em uma área que abrangeu 48,75 ha de cobertura florestal primária. Como proceder na escolha das espécies a serem replantadas, de forma a considerar não tão somente a disponibilidade de sementes num dado momento, mas também, por exemplo, em fatores inerentes à fitossociologia horizontal das espécies presentes na floresta primitiva, anterior à supressão da vegetação para a lavra do minério? Neste sentido, são gerados índices estatísticos para, entre outras funções, auxiliar o silvicultor na tomada de decisões a esse respeito.
COMPARAÇÃO ENTRE OS ÍNDICES FITOSSOCIOLÓGICOS
Uma comparação entre os índices IFH e IVI foi efetuada com o intuito de avaliar qual deles trazia mais informações acerca das espécies e das associações existentes na floresta ombrófila submontana densa, a qual apresentou uma alta abundância e riqueza de espécies.
O ranqueamento das espécies, feito de forma uniforme para os índices, permitiu visualizar que, para os índices IFH e IVI, foram classificadas na categoria de prioridade A (alta) 11 e três espécies, na categoria B (intermediária) 54 e três espécies, e na categoria C (baixa) 731 e 790 espécies, respectivamente para os índices IFH e IVI (Tabela 2). Todas as espécies classificadas como de alta e média prioridade, pelo índice IFH, encontram-se relacionadas na Tabela 3, incluindo o IVI e suas respectivas variáveis absolutas de abundância, frequência e dominância.
O índice IVI classificou apenas três espécies como de alta prioridade (Geissospermum sericeum, Eschweilera coriacea e Rinorea riana), enquanto o índice IFH hierarquizou 11 espécies (Tabela 3). Observou-se que o índice IFH conseguiu captar, na categoria de prioridade alta, três espécies do gênero Eschweilera e duas das principais famílias (Sapotaceae e Lecythidaceae). Considerando-se, então, que os resultados referentes às espécies de alta prioridade não foram suficientes para explicar as espécies prioritárias da fitofisionomia estudada, as categorias A e B foram agruapadas em uma única, quando então foram observadas 65 espécies para o IFH e seis espécies para o IVI nesta nova categoria (Tabela 2).
Mesmo com este procedimento, o IVI não incluiu nenhuma espécie de Pouteria e Protium, importantes gêneros na caracterização da florística de florestas amazônicas, sobretudo no local deste estudo. Por considerar tão somente a soma aritmética das três variáveis que o formam, todas com o mesmo peso, este índice não conseguiu transmitir uma análise consistente da composição florística da floresta através das seis espécies-chave por ele classificadas como de prioridade ecológica alta e intermediária.
O IVI, quando proposto por Curtis & McIntosh (1951), foi desenvolvido para florestas temperadas do sudeste do Wisconsin, nos Estados Unidos, com poucas espécies (25). Ao se generalizar o seu emprego para todas as demais fitofisionomias, é de se esperar que tais inconsistências sejam captadas. Uma floresta temperada com 25 espécies é completamente diferente de uma floresta tropical amazônica com 796 espécies, distribuídas em 58 famílias. Um índice eficaz deve transmitir ao tomador de decisão uma visão realista e mais sumarizada possível do todo, de tal forma que contenha os principais componentes avaliados, traduzidos em algumas dezenas de espécies-chave da fitofisionomia, assim como deve apresentar as principais famílias. O IVI não permitiu visualizar nem uma dezena de espécies-chave, nem as famílias mais características e tampouco as associações de gêneros descritas por diversos autores, para diferentes locais da Amazônia.
O IFH, obtido por meio de análise multivariada, pelo método de componentes principais, considerando uma variável dummy (qualitativa) para cada uma das variáveis incluídas no modelo, conseguiu traduzir com muito mais fidelidade a sumarização da composição florística predominante da floresta analisada. Consequentemente, gerou uma melhor compreensão da complexa composição florística representada pelas 796 espécies registradas. As 65 espécies classificadas como de prioridade alta e intermediária (8,17% do total) envolveram as famílias e as associações de gêneros mais características da fitocenose estudada. As famílias com maior riqueza e abundância, relatadas em diversos estudos para diferentes locais da Amazônia, foram captadas por este índice, destacando-se Sapotaceae (14 espécies), Leguminosae sensu lato (9), Annonaceae (5) e Burseraceae, Chrysobalanaceae, Euphorbiaceae e Mimosaceae (4 de cada), além de Fabaceae, Lecythidaceae, Myristicaceae e Violaceae (3 de cada). As associações de gêneros intrinsecamente presentes nas diversas facies da floresta amazônica, como Pouteria, Escheweilera e Protium (respectivamente 10, 3 e 3 espécies), também foram captadas pelo índice. Finalmente, as 65 espécies ranqueadas pelo IFH como de prioridade ecológica alta e intermediária, com ampla distribuição na área (12.122 indivíduos em 48,75 ha da amostra), indicam que este índice mostrou-se eficiente para uma análise da fitossociologia horizontal das espécies (Tabela 3).
Observou-se que os resultados do IFH atenderam mais amplamente as recomendações da Resolução SMA 47, de 21/11/2003, do estado de São Paulo (SMA, 2003), que preconiza para os trabalhos de restauração florestal, em áreas originalmente cobertas por florestas, um número mínimo de 80 espécies e também ao Decreto no. 31.594, de 27/01/2010, do estado do Pará (IDEFLOR, 2010), que recomenda a utilização do maior número possível de espécies nativas de ocorrência regional para o plantio de mudas com fins de recomposição da Reserva Legal (80% da área total do imóvel na Amazônia).
No entanto, há de se frisar que todas as espécies são únicas e relacionadas entre si no complexo ecossistema florestal amazônico. Ao se procurar caracterizar as espécies-chave de um determinado ecossistema, não se pode excluir da análise as demais, pois, como se sabe, entre 17% e 33% das espécies ocorrem com apenas um indivíduo na totalidade dos levantamentos executados em florestas primárias na Amazônia (Salomão et al., 2007). Neste trabalho, foi observada a ocorrência de 132 espécies (ou 16,6% do total) que ocorreram com apenas um indivíduo, podendo ser consideradas como localmente raras; outras 81 espécies (10,2%) ocorreram com apenas dois indivíduos.
CONCLUSÃO
Os testes estatísticos validaram a amostra de dados para o emprego da técnica de análise multivariada e, portanto, para a construção do índice de fitossociologia horizontal (IFH).
Para a análise da fitossociologia horizontal da floresta ombrófila submontana densa, o índice de fitossociologia horizontal com variável dummy (IFH), obtido por meio da análise multivariada, apresentou maior eficiência do que o índice de valor de importância (IVI) na seleção de espécies-chave para a restauração florestal de áreas mineradas.
Considerando as espécies de alta e intermediária prioridade como sendo as prioritárias para o plantio nas áreas anuais de restauração florestal, o IFH proposto conseguiu captar um número de espécies-chave 10,8 vezes superior ao IVI.
A indicação do índice proposto neste trabalho como metodologia alternativa, em função do rigor científico, deve merecer aplicações em outras comunidades vegetais para efeito de sua validação.
Estes índices podem ser de grande contribuição para as propostas que visam a recuperação de áreas degradadas pelas atividades agropecuárias e minerárias, por meio da indicação das espécies-chave para o plantio, visando restaurar a paisagem florestal.
Sendo as unidades de conservação, inclusive a FLONA Saracá-Taquera/ICMBio, um patrimônio da sociedade brasileira, constituindo-se em um rico acervo de recursos naturais e biológicos, a sua restauração deve atender aos aspectos legais vigentes e também a princípios científicos rígidos, no sentido de recompor a paisagem florestal, tornando-a o mais próximo possível da originalidade anterior às atividades econômicas ali desenvolvidas.
AGRADECIMENTOS
À Mineração Rio do Norte (MRN), financiadora deste estudo, e a seus funcionários, em especial, ao Ademar Cavalcanti; ao José Risonei Assis da Silva e demais servidores da FLONA Saracá-Taquera/lCMBio; ao Nélson A. Rosa, do Museu Paraense Emílio Goeldi, pelas identificações botânicas e ao Hans ter Steege, da Universidade de Utrecht, Holanda, pela completa revisão das identificações para o sistema APG lll, expressamos nossos sinceros agradecimentos. Ao trabalho anônimo dos revisores, que contribuíram para um melhor aperfeiçoamento do artigo, manifestamos também nossa gratidão.
REFERÊNCIAS
BLACK, G. A., T. H. DOBZHANSKY & C. PAVAN, 1950. Some attemps to estimate species diversity and population density of trees in Amazonian forests. Botanical Gazette 111(4): 413-425.
CAIN, S. A., G. M. O. CASTRO, J. M. PIRES & N. L. SILVA, 1956. Application of some phytosociological techniques to Brazilian rain forest. American Journal of Botany 43: 911-941.
CAMPOS, J. C. C. & H. G. LEITE, 2002. Mensuração florestal: perguntas e respostas: 1-407. Editora UFV Viçosa.
CARPANEZZI, A. A., L. G. S. COSTA, P. Y. KAGEYAMA & C. F. A. CASTRO, 1990. Espécies pioneiras para recuperação de áreas degradadas: a observação de laboratórios naturais. Anais do Congresso Florestal Brasileiro 6: 216-221.
CURTIS, J. T. & R. P. MCINTOSH, 1951. An upland forest continuum in the prairie-forest border region of Wisconsin. Ecology 32(3): 476-496.
DAVIS, T. A. W. & P. W. RICHARDS, 1934. The vegetation of Moraballi Creek, British Guiana: an ecological study of a limited area of tropical rain forest ll. Journal of Ecology 22: 106-155.
DILLON, W. R. & M. GOLDSTEIN, 1984. Multivariate analysis: methods and applications: 1-415. John Wiley & Sons, New York.
GAMA, Z. J. C., A. C. SANTANA, F. A. T. MENDES & A. S. KHAN, 2007. índice de desempenho competitivo das empresas de móveis da Região Metropolitana de Belém. Revista de Economia e Agronegócio 5(1): 127-160.
GAUCH, H. G., 1982. Multivariate analysis in community ecology: 1-298. Cambridge University Press, Cambridge.
GOODALL, D. W., 1954. Objective methods for the classification of vegetation. III. An essay in the use of factor analysis. Australian Journal of Botany 2(3): 304-324.
HAIR, J. F., R. E. ANDERSON, R. L. TATHAM & W. C. BLACK, 2005. Análise multivariada de dados. 5. ed.: 1-258. Bookman, Porto Alegre.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA (IBGE), 1992. Manual Técnico da Vegetação Brasileira: 1-92. IBGE (Série Manuais Técnicos em Geociências, 1), Rio de Janeiro.
INSTITUTO DE DESENVOLVIMENTO FLORESTAL DO ESTADO DO PARÁ (IDEFLOR), 2010. Instrução Normativa No 02/2010. Disponível em: <http://www.ideflor.pa.gov.br/files/u3/IN02-2010.pdf>. Acesso em: 18 outubro 2010.
JOHNSON, R. A. & D. W. WICHERN, 1992. Applied multivariate statistical analysis: 1-390. Prentice- Hall, New Jersey.
JOHNSON, R. A. & D. W. WICHERN, 2007. Applied multivariate statistical analysis. 6. ed.: 1-800. Prentice-Hall, New Jersey.
MARGALEF, R., 1972. Homage to Evelyn Hutchinson or why is there an upper lilit to diversity. Transactions of the Illinois State Academy of Science 44: 221-235.
MINGOTI, S. A., 2005. Análise de dados através de métodos de estatística multivariada: uma abordagem aplicada: 1-450. Editora UFMG, Belo Horizonte.
MOSCATELLI, M., C. DE'CARLI & J. R. ALMEIDA, 1993. Legalidade teórica e realidade prática na defesa de manguezais: estudo de caso em Angra dos Reis, Rio de Janeiro. Anais do Simpósio de Ecossistemas da Costa Brasileira: Subsídios a um Gerenciamento Ambiental 3: 93-98.
OLIVEIRA, A. N. & I. L. AMARAL, 2004. Florística e fitossociologia de uma floresta de vertente na Amazônia Central, Amazonas, Brasil. Acta Amazonica 34(1): 21-34.
OLIVEIRA-FILHO, A. T & J. A. RATTER, 1995. A study ofthe origin of central Brazilian forests by the analysis of plant species distribution patterns. Edinburgh Journal of Botany 52(2): 141-194.
PIELOU, E. C., 1975. Ecological diversity: 1-403. Wiley-Interscience, New York.
PIELOU, E. C., 1984. The interpretation of ecological data: a primer on classification and ordination: 1-288. Wiley-1nterscience, New York.
PIRES, J. M., T. H. DOBZHANSKY & G. A. BLACK, 1953. An estimate of the number of trees in an Amazonian forest community. Botanical Gazette 114(4): 467-477.
PRANCE, G. T., W. A. RODRIGUES & M. F. SILVA, 1976. Inventário florestal de um hectare de mata de terra firme, km 30 da Estrada Manaus-Itacoatiara. Acta Amazônica 6(1): 9-35.
QUEIROZ, W. T. & A. V. BARROS, 1998. Inventário floresta de 3.097 ha da Floresta Nacional de Saracá-Tacuera - Município de Oriximiná - Pará: 1-312. Mineração Rio do Norte, Faculdade de Ciências Agrárias do Pará (Relatório Técnico), Belém.
RANKIN-DE-MERONA, J. M., G. T. PRANCE, R. W. HUTCHINGS, M. F. SILVA, W. A. RODRIGUES & M. E. UEHLING, 1992. Preliminary results of a large-scale tree inventory of upland rain forest in the Central Amazon. Acta Amazonica 22(4): 493-534.
RAUNKIAER, C., 1934. The life forms of plants and statistical geography. The Clarendon Press, Oxford.
RODRIGUES, R. R., R. A. F. LIMA, S. GANDOLFI & A. G. NAVE, 2009. On the restoration of high diversity forests: 30 years of experience in the Brazilian Atlantic Forest. Biological Conservation 142: 1242-1251.
ROLLET, B., 1993. Tree populations in natural tropical rain forest. Bois et Forests des Tropiques 236(2): 43-55.
SALOMÃO, R. P., I. C. G. VIEIRA, C. SUEMITSU, N. A. ROSA, S. S. ALMEIDA, D. D. AMARAL & M. P. M. MENEZES, 2007. As florestas de Belo Monte na grande curva do rio Xingu, Amazônia Oriental. Boletim do Museu Paraense Emílio Goeldi. Ciências Naturais 2(3): 57-153.
SANQUETTA, C. R., L. F. WATZLAWICK, A. P. D. CÔRTE, L. A. V. FERNANDES & J. D. P. SIQUEIRA, 2009. Inventários florestais: planejamento e execução: 2: 1-316. Multi-Grafhic Editora, Curitiba.
SANTANA, A. C., 2005. Elementos de economia, agronegócio e desenvolvimento local: 133-142. GTZ; TUD; UFRA (Série Acadêmica, 1), Belém.
SANTANA, A. C., 2007. Índice de desempenho competitivo das empresas de polpa de frutas do Estado do Pará. Revista de Economia e Sociologia Rural 45(3): 749-775.
SANTANA, A. C., 2008. Índice de desempenho competitivo das empresas de polpa de frutas do Estado do Pará. In: A. S. CORDEIRO, D. F. CARVALHO & F. A. T. MENDES (Orgs.): Análise sistêmica da fruticultura paraense: organização, mercado e competitividade empresarial. 1. ed.: 115-143. Banco da Amazônia, Belém.
SANTOS, R. M., F. A. VIEIRA, M. FAGUNDES, Y. R. F. NUNES & E. GUSMÃO, 2007. Riqueza e similaridade florística de oito remanescentes florestais no norte de Minas Gerais, Brasil. Revista Árvore 31(1): 135-144.
SECRETARIA DE MEIO AMBIENTE DO ESTADO DE SÃO PAULO (SMA), 2003. Resolução SMA 43, de 21/11/2003. Diponível em: <http://licenciamento.cetesb.sp.gov.br/legislacao/estadual/resolucoes/2010_Res_SMA_43.pdf>. Acesso em: 4 junho 2010.
SHANNON, C. E. & W. WIENER, 1949. The mathematical theory of communication: 1-117. University of Illinois Press, Urbana.
SILVA, A. F. & G. J. SHEPHERD, 1986. Comparações florísticas entre algumas matas em floresta densa da bacia do Rio Juruá. Boletim do Museu Paraense Emílio Goeldi, série Botânica 8(2): 203-258.
SILVA, A. S. L., P. L. B. LISBOA & U. N. MACIEL, 1992. Diversidade florística e estrutura em floresta densa da bacia do Rio Juruá. Boletim do Museu Paraense Emílio Goeldi, série Botânica 8(2): 203-258.
SIMPSON, E. H., 1949. Measurement of diversity. Nature 163: 688-697.
SOARES, C. P. B., F. PAULA NETO & A. L. SOUZA, 2006. Dendrometria e inventário florestal: 1-276. Editora UFV, Viçosa.
SOCIETY FOR ECOLOGIAL RESTORATION (SER), 2004. Princípios da SER International sobre a restauração ecológica. Society for Ecological Restoration International, Tucson. Disponível em: <http://www.ser.org>. Acesso em: 4 outubro 2010.
STELLA, A., 2002. Seleção de especies arboreas tropicais prioritárias para a conservação: o caso da Estação Ecologica de Caetetus. Dissertação (Mestrado em Biologia Vegetal) - Universidade de Campinas, Campinas. Disponível em: <http://www.bibliotecadigital.unicamp.br/document/?code=vtls0002676 91&fd=y>. Acesso em: 4 outubro 2010.
SUPERINTENDÊNCIA DO DESENVOLVIMENTO DA AMAZÔNIA (SUDAM), 1973. Levantamentos florestais realizados pela missão FAO na Amazônia: 1-798. Superintendência do Desenvolvimento da Amazônia, Belém.
TOLEDO, L. O., L. H. C. ANJOS, W. H. COUTO, J. R. CORREIA, M. G. PEREIRA & M. E. F. CORREIA, 2009. Análise multivariada de atributos pedológicos e fitossociológicos aplicada na caracterização de ambientes de cerrado no norte de Minas Gerais. Revista Árvore 33(5): 957-968.
VALENCIA, R., H. BALSLEV & G. C. PAZ Y MINO, 1994. High tree alpha-diversity in Amazonian Ecuador. Biodiversity and Conservation 3(1): 21-28.
VIANA, V. M., 1990. Biologia e manejo de fragmentos de florestas naturais. Anais do Congresso Florestal Brasileiro 6: 219-221.
 Autor de correspondência:
Autor de correspondência:
Rafael de Paiva Salomão.
Museu Paraense Emílio Goeldi/Ministério da Ciência,
Tecnologia e Inovação.
Coordenação de Botânica.
Av. Perimetral, 1901 - Terra Firme. Belém, PA, Brasil.
CEP 66017-970
(salomao@museu-goeldi.br).
Recebido em 05/10/2010
Aprovado em 11/04/2012
Responsabilidade editorial: Hilton Tulio Costi
