










Serviços Personalizados
Journal

Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Curriculum ScienTI
Curriculum ScienTI Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Pan-Amazônica de Saúde
versão impressa ISSN 2176-6215versão On-line ISSN 2176-6223
Rev Pan-Amaz Saude v.1 n.1 Ananindeua mar. 2010
http://dx.doi.org/10.5123/S2176-62232010000100011
Estudio experimental sobre la patogenicidad del Virus Ilhéus en hámsteres dorados (Mesocricetus auratus)
Raimunda do Socorro da Silva AzevedoI; Vera Lúcia Reis de Souza BarrosII; Lívia Carício MartinsI; Ana Cecília Ribeiro CruzIII; Sueli Guerreiro RodriguesI; Pedro Fernando da Costa VasconcelosIII
ISeção de Arbovirologia
e Febres Hemorrágicas, Instituto Evandro Chagas/SVS/MS, Ananindeua,
Pará, Brasil
IISeção de Patologia, Instituto Evandro Chagas/SVS/MS,
Ananindeua, Pará, Brasil
IIISeção de Arbovirologia e Febres Hemorrágicas,
Instituto Evandro Chagas/SVS/MS, Ananindeua, Pará, Brasil. Universidade
do Estado do Pará, Belém, Pará, Brasil
Endereço para
correspondência
Correspondence
Dirección para correspondencia
Título original: Estudo experimental sobre a patogenicidade do Vírus Ilhéus em hamsters dourados (Mesocricetus auratus). Traducido por: Rocio Tamara (resumen) y Lota Moncada (artículo)
RESUMEN
Con el fin de investigar la patogenicidad del Flavivirus Ilheus (VILH), se inoculó por vía intraperitoneal (VIP) 9,8 DL50 de suspensión viral en hámsteres dorados jóvenes (Mesocricetus auratus), y se obtuvieron diariamente bajo anestesia muestras de sueros y órganos (cerebro, hígado, corazón, bazo, riñones y pulmones) de animales infectados y controles no infectados. Durante el experimento se determinó el título viral del VILH en el suero y órganos infectados, en ratones recién nacidos. Además, se realizó en los sueros la detección de antígeno y los niveles de anticuerpos, a través de pruebas de fijación del complemento y de inhibición de la hemoaglutinación. El examen histopatológico por HE y la detección de antígenos virales por inmunohistoquímica (IHQ), se llevó a cabo en los tejidos de los animales. La dosis inoculada ocasionó la muerte de los animales por encefalitis en el séptimo día tras la inoculación. Todos los órganos estudiados mostraron cambios en los tejidos detectables por histopatología. Se detectó por IHQ una presencia masiva del antígeno del virus en el cerebro y en menor medida en el hígado, bazo y riñones, aunque en estos órganos la presencia de antígeno viral fue transitoria y leve, lo que corroboró con los títulos virales obtenidos en estos órganos. No fueron encontrados antígenos virales en el corazón y en los pulmones, lo que sugiere que los títulos (DL50) observados en estos órganos, durante la titulación viral en los ratones, son el resultado de la presencia de VILH en el torrente sanguíneo (viremia). Los resultados de este estudio refuerzan el importante y conocido neurotropismo del VILH.
Palabras clave: Virus Ilhéus; Flavivirus; Encefalitis Arbovirus; Virulencia; Modelos Animales.
INTRODUCCIÓN
El Virus Ilhéus (VILH) pertenece al género Flavivirus de la familia Flaviviridae, y ha sido inserido al grupo Ntaya virus7. Sin embargo, estudio molecular reciente demostró de forma vehemente que el VILH, junto con el ROCV, representan un linaje genético distinto al grupo Ntaya virus10. Luego del primer aislamiento viral, obtenido en 1944, innumerables otros aislamientos, de pacientes febriles, de mosquitos y de una gran variedad de animales, particularmente aves silvestres y murciélagos, se obtuvieron en Brasil, Colombia, Panamá, Argentina y en Trinidad8,11,12,19,23. En la Amazonia brasileña el virus ha sido aislado de pacientes febriles, de monos centinelas, de murciélagos y de diversas especies de mosquitos, principalmente el Psorophora ferox, considerado su principal vetar14,19,20,22. Aves silvestres han sido implicadas como probables hospederos vertebrados del virus, sin embargo, también han sido encontrados anticuerpos, o aislado el virus en otros vertebrados, como roedores, marsupiales, desdentados, murciélagos y monos4,6,8.
La enfermedad en humanos usualmente se encuentra en casos esporádicos asociados a exposición en regiones de floresta, y el cuadro clinico varia desde infecciones asintomáticas hasta encefalitis. La enfermedad febril, responsable por la mayoria de los casos reconocidos, se inicia súbitamente, con fiebre moderada o elevada, cefalea, escalofríos, fotofobia, artralgia, mialgias y astenia, evolucionando en un promedio de 3 a 5 dias, con recuperación completa y sin secuelas. El pequeño número de casos clinicos diagnosticados contrasta con la elevada prevalencia de anticuerpos para el agente, lo que sugiere que gran parte de las infecciones sea no aparente o oligosintomática. Otro factor relevante es su corto periodo de incubación. Durante la observación de ocho casos agudos, con aislamiento del VILH, ocurridos en Brasil, en Trinidad, en Argentina, en Colombia y en Panamá, en un paciente fueron relatadas alteraciones del sistema nervioso central que llevaron a la sospecha de encefalitis13.
Pocas informaciones sobre estudios relacionados con la patogenicidad del VILH están disponibles. Sin embargo, se sabe que estudios experimentales con el virus en animales de pequeño porte, como ratones y otros roedores, bien como en primates no humanos, produjeron viremia8,9. Experimentos realizados por Tesh y colaboradores21 y Xiao y colaboradores24,25 en hámsteres dorados (Mesocricetus auratus), para estudiar la patogénesis de flavivirus, demostraron gran susceptibilidad de este modelo experimental y, como representan excelente modelo alternativo, en sustitución a primates no humanos, decidimos realizar experimentos via intraperitoneal (VIP) con el VILH en esos animales.
MATERIAL Y MÉTODOS
La cepa del VILH BeH 7445 utilizada en este estudio es proveniente de suspensión de cerebros de ratones albinos Swiss infectados luego de 21 pasajes, y mantenidos a - 70o C, sin pasaje anterior en hámster. Esta cepa viral fue aislada en 1959 a partir de la sangre de un paciente febril, procedente del distrito de Caraparu, localizado en el Municipio de Santa Isabel de Pará, en el Estado de Pará, Brasil2.
La inoculación se preparó a partir de cerebros de ratones albinos infectados con la cepa del VILH, en solución salina tamponada con fosfato pH 7,4 conteniendo albúmina bovina 0,75% y antibióticos (100 Ul/mL de penicilina y 100 µg/mL de estreptomicina), y mantenido a temperaturas inferiores a 8o C. Se inocularon 0,1 mL de la suspensión en veinte hámsteres machos, de 4 a 5 semanas de edad, por VIP, reservándose diez animales para control negativo (no inoculados). Con la finalidad de detectar la dosis aplicada, la solución viral también fue inoculada en diluciones seriadas 10-1 a 10-12 en ratones con 2 a 3 dias de edad. El titulo viral se calculó por el método de Reed y Muench16 y expresado como DL50/0,02 mL. A cada 24 horas post inoculación, fueron anestesiados y sacrificados dos hámsteres infectados y uno del grupo de control, para sangria y recolección de órganos (cerebro, higado, corazón, bazo, riñón y pulmón). Una alícuota del suero fue guardada a -70° C, para estudios sobre la viremia y detección de antigenos, y otra a -20o C para detección de anticuerpos. Los fragmentos de visceras recolectados fueron divididos en dos partes, una para congelamiento (-70° C) destinada a la detección de antigenos y titulación viral en los tejidos, y la otra fue sumergida en solución de formalina tamponada a 10%, para uso en examen histopatológico e inmunohistoquimico. Los especimenes recogidos fueron, por lo tanto, utilizados para determinación de viremia por la titulación en ratones recién-nacidos, detección de antigenos por el test de fijación del complemento (FC) e inmunohistoquimica (IHQ), detección de anticuerpos por los tests de FC e inhibición de la hemoaglutinación (IH) y examen tecidual (histopatologia).
Con el fin de detectar la presencia del VILH, bien como de analizar la curva virémica producida por el virus en los especimenes biológicos obtenidos del ensayo experimental, la titulación viral se realizó separadamente, a partir de sueros y fragmentos de órganos con preparo de suspensión de macerado a 10% en PBS conteniendo albúmina bovina y antibióticos. El sobrenadante fue utilizado para preparar diluciones seriadas de 10-1 a 10-12 que fueron inoculadas a un grupo de seis ratones recién-nacidos, administrando 0,02 mL por via intracerebral a cada animal. Los titulos también fueron calculados por el método de Reed y Muench16.
O test de FC fue utilizado para la detección de antigenos y anticuerpos del VILH. Los tests se realizaron conforme la técnica modificada de Fulton y Dumbell5 y adaptada para microplaca por Shope y Sather17, en diluciones en serie de 2x (1:8 a 1:128 para las fuentes de antigeno y de 1:8 a 1:64 para las fuentes de anticuerpos), utilizando dos unidades de complemento de cobaya, antigeno (usado como control del test) y fluido ascitico hiperinmune del ILH (suero anti-VILH) y del Bunyavirus Tacaiuma, cepa viral BeAN 73 (suero anti-TCM) y hematies de carnero sensibilizadas. La lectura se hizo de acuerdo a los porcentajes de hemólisis observados, y los titulos positivos fueron registrados como la más alta dilución con un porcentual de hemólisis de hasta 25%.
Para el estudio de los anticuerpos IH fue utilizada la técnica de Clarke y Casals3, adaptada para microplacas usando diluciones de 1:20 até 1:1280 de los sueros, contra cuatro unidades del antigeno del VILH y del control negativo (virus TCM).
Los fragmentos de visceras fijados en formalina tamponada a 10% fueron encaminados y procesados para microscopia óptica. La técnica utilizada fue la de Hematoxilina - Eosina15. Los órganos estudiados fueron semicuantificados en una escala variando de 0 a 3, siendo: 0= ausencia de lesión; 1= lesión ligera; 2= lesión moderada y 3= lesión intensa25. De todos los fragmentos de visceras de los animales infectados y de los animales control, ya emblocados en parafina, fueron retirados cortes de 5 µM y fijados en láminas Super-frost®/Plus (A. Dalgger & Company). Los cortes fueron teñidos, de acuerdo a los procedimientos descritos por Barros1, utilizando fluido ascitico hiperinmune del VILH.
RESULTADOS
El título DL50/0,02 mL de la cepa BeH 7445 del VILH inoculada VIP en hámsteres jóvenes fue de 9,8. La colecta de sangre (para obtención de suero) y de los fragmentos de visceras sucedió durante siente días, ya que después de ese período, todos los animales evolucionaron hacia el óbito. En el experimento encontramos, en promedio, títulos virales relativamente bajos, sin embargo, detectables en las primeras 24 h, siendo que, en el suero colectado en el segunda día post inoculación (p.i.) fue encontrado el título más elevado (DL50/0,02 mL = 5,5). En fragmentos de hígado, corazón y bazo observamos que el pico máximo se dio con 24 h p.i. En fragmentos de cerebro, riñón y pulmón observamos que el pico máximo sucedió en el quinta, tercer y segunda días luego de la infección, respectivamente (Figura 1).
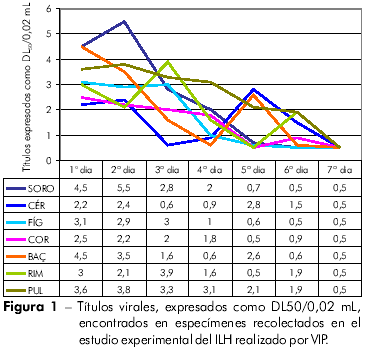
En este estudio con el VILH, se demostró que, en las muestras de suero recogidas, la viremia fue detectable en las primeras 24 h p.i., con duración de cuatro días y pico máximo al segunda dia p.i. En los fragmentos de vísceras, la presencia del VILH se detectó desde las primeras 24 h p.i., con duración variando de cuatro a seis días. Interesante observar el comportamiento de la titulación viral en el cerebro, que fue claramente de evolución bifásica. De hecho, en los dos primeros días post inoculación, se observó el título virémico DL50/0,02 mL discretamente mayor que 2. A partir de entonces, al tercera e cuarta días p.i. hubo un evidente descenso para 0,6 e 0,9 DL50/0,02 mL, respectivamente. Al quinta día p.i. el título llegó al máximo (2,8 DL50/0,02 mL), manteniéndose con 1,5 DL50/0,02 mL al sexto día p.i. para ya no ser más detectado al sétimo día, como observado para todos los especímenes estudiados (Figura 1).
Los estudios realizados para detección de anticuerpos demostraron el aparecimiento de anticuerpos fijadores del complemento a partir del cuarto día p.i., bien como de anticuerpos inhibidores de la hemoaglutinación a partir del quinta día luego de la infección experimental (cuadro 1). A pesar del aparecimiento de los anticuerpos, todos los animales inoculados, como dicho antes, evolucionaron al óbito.

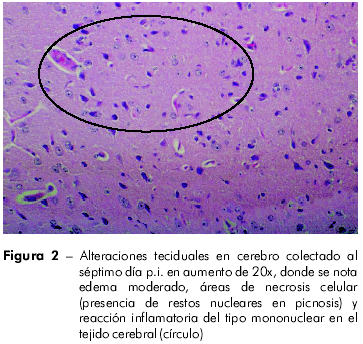
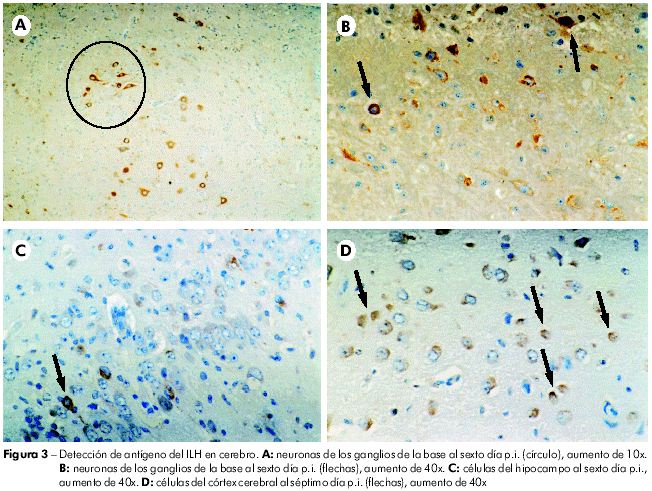
El estudio histopatológico (HE) reveló en el cérebro leptomeningitis, edema leve y congestión, que se tornan más intensos a partir del quinta día p.i. En el córtex y sustancia blanca se percibieron, del primero ao cuarto dia p.i., congestión de leve a moderada intensidad, y edema intenso, más visible en el espacio perivascular, acompañado de reacción inflamatoria (presencia de células aisladas o en pequeños grupos, mostrando citoplasma retraído, acidófilo y núcleo picnótico). Del quinta ao séptimo día p.i., el edema se presentó moderado en el espacio pericelular, habiendo también aumento Del número de áreas con neuronas acidófilas o en necrosis celular, documentada por la presencia de restos nucleares picnóticos (Figura 2). La IHQ demostró inmunomarcación, a partir del 5º día p.i. (120 h p.i.), inicialmente en gânglios de la base, extendiéndose en el correr de los demás dias post-infección al hipocampo y al córtex cerebral (Figura 3 ).
En el hígado, la red trabecular se mantuvo del inicio al fin del experimento. Al primero día p.i. se percibió una tumefacción celular intensa, principalmente en las zonas dos (medio-zonal) y tres (espacio puerta) del acino hepático, asociada a la acidofilia citoplasmática en células parenquimales aisladas, además de la presencia de células inflamatorias raras en los sinusoides. No segundo día p.i., además de la tumefacción celular, se nota retracción acidofílica del citoplasma de algunas células que presentaron también núcleo picnótico, además del surgimiento de estructuras corpusculares, con las características de los corpúsculos de Councilman-Rocha Lima, de distribución esparcida (células en apoptosis). Los sinusoides quedaron realzados y exhibieron hipertrofia e hiperplasia de células de Kupffer y moderada reacción inflamatoria representada principalmente por mononucleares. Ya en los tractos portales la reacción inflamatoria fue leve. AL tercer e cuarto días p.i. la lesión se presenta con moderada intensidad, caracterizándose por la presencia de necrosis celular focal, esparcida, en el parénquima, mayor número de corpúsculos tipo Councilman, además de la moderada reacción inflamatoria intralobular y en los tractos portales. Al quinto día p.i., además de las alteraciones observadas anteriormente, se percibe congestión focal en los sinusoides, sin embargo, ya estaba presente la binucleación del hepatócito, una típica señal de instalación del proceso regenerativo. Del sexto al séptimo día p.i. se notó regresión de las lesiones y acentuación del proceso regenerativo hepático. La IHQ reveló hepatócitos marcados apenas al segundo día (48 h) post-infección, no encontrándose antígenos virales en los demás días del experimento (Figura 4).
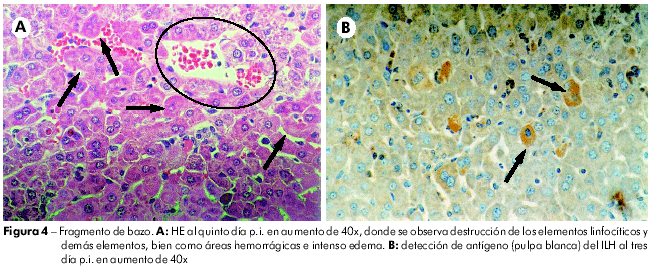
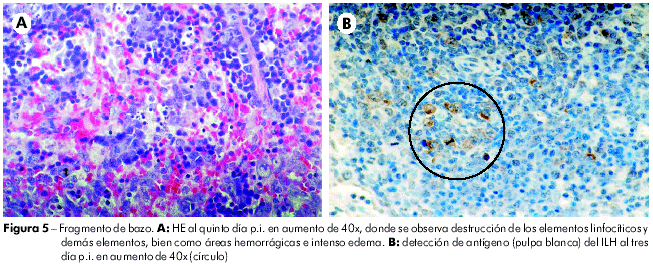
En el bazo, al primero día p.i., se notó que la pulpa blanca presentaba hiperplasia folicular y la pulpa roja ya exhibía cierto grado de congestión. Del segundo Del quinto días p.i. se nota presencia de necro-apoptosis de los elementos linfocíticos, en la pulpa blanca y en la pulpa roja, y que se acompañaban del surgimiento de macrófagos conteniendo cuerpos teñibles, de tal manera que al tercero día p.i. ya no fueron identificados más elementos linfáticos íntegros, apenas cuerpos apoptóticos y polvo nuclear; y, al quinto día, ya se observaba, en medio a restos celulares, la presencia de centroblastos e inmunoblastos, además de cierto número de plasmocitos. Con el paso de los días hubo una gradual elevación de esas células. En la pulpa roja se notó, además, moderada congestión al segundo día, que se tornó intensa, siendo acompañada de áreas hemorrágicas en los tercero, cuarto, y quinto días p.i. Del sexto al séptimo día p.i. se observó persistencia de restos celulares de forma leve, con disminución del proceso congestivo e intensa proliferación de centroblastos e inmunoblastos, indicando fase de regeneración tecidual. También se observó la presencia de plasmocitos. En la pulpa blanca, se detectó inmunomarcación de antígenos virales al tercero día (72 h) p.i. Como observado en el hígado, los demás días del experimento no se observó presencia de antígenos virales en las células esplénicas (Figura 5).
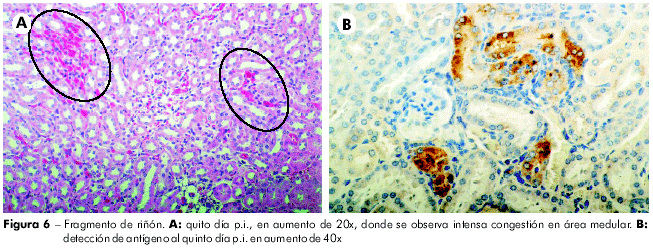
En los riñones, al primero día p.i., se percibe apenas congestión focal intertubular. Del segundo al quinto días p.i. los glomérulos se mantuvieron preservados, sin embargo, apareció tumefacción leve en células tubulares, observándose que la congestión en el estroma intertubular se torna intensa con el avance del experimento, siendo más evidente en la porción medular. Ya al sexto y séptimo días p.i., glomérulos y túbulos están preservados, sin embargo, se observa congestión intersticial leve, bastante más evidente en la porción cortical renal. En la IHQ se observa la presencia del antígeno viral en túbulos renales en el cuarto e quinto días p.i. Los demás días del experimento no se detecta la presencia de antígenos virales (Figura 6).
En el corazón, las alteraciones histológicas se percibieron a partir del segundo día p.i., caracterizándose por edema intersticial leve e hiperemia difusa, bastante más evidentes en el miocardio. Tales lesiones se extendieron hasta el quinto día p.i. En los sexto y séptimo días p.i., ya no se percibían alteraciones morfológicas en el corazón. En los pulmones, los dos primeros días p.i. no se perciben alteraciones histológicas. Del tercero al quinto días p.i. se observan áreas mostrando colapso alveolar, además de moderada congestión de pared de alvéolos y hemorragia intra-alveolar de ligera a moderada intensidad. Al sexto y séptimo días p.i., se nota apenas congestión leve de paredes alveolares. En ambas vísceras examinadas (corazón y pulmón) no se encontró inmunomarcación en ninguno de los días del experimento.
DISCUSIÓN
Aunque haya sido utilizada en el experimento una cepa viral que contenía 21 pasajes en ratones recién-nacidos, la misma nunca había sido inoculada en hámsteres. A pesar de la posibilidad de que haya habido alteración durante los sucesivos pasajes VIC en ratones, fue interesante observar que los animales experimentados desarrollaron bajos títulos virales en todos los órganos, siendo más duraderos en los pulmones que en el suero o en cualquier otra víscera. ¿Será que los animales murieron por lesión pulmonar? La histopatología y la IHQ, sin embargo, no confirmaron esta posibilidad, ya que el pulmón fue el órgano con menores alteraciones observadas en el estudio histopatológico, e, además, antígenos virales no fueron detectados por la IHQ en este órgano. No obstante, se observó por HE la leptomeningitis progresiva y necrosis tisular en el cerebro, hallazgos que, corroborados por la detección de antígeno viral por IHQ a partir del quinto día p.i en el cerebro, demuestran que el VILH determinó una meningoencefalitis en esos animale (Figura 3). En los fragmentos de hígado, bazo y riñón recogidos, en los que los títulos fueron bajos, se detectaron antígenos del VILH por IHQ al segundo día p.i. en el hígado, al tercero día p.i. en el bazo y al cuarto y quinto días p.i. en el riñón, demostrando el tropismo del VILH para esos órganos, siendo destacable la capacidad del VILH de inducir producción de antígenos en esos órganos en diferentes fases evolutivas de la infección (Figuras 4, 5 e 6). En fragmentos de pulmón y corazón, a pesar de haber demostrado títulos virales, bien como discretas alteraciones teciduales, no se detectó la presencia de antígeno viral por IHQ. Esto puede significar que los títulos virales, así como las lesiones teciduales encontradas son consecuencia del pasaje del virus por esos órganos a través de la sangre, como mencionado anteriormente.
Comparando el título viral con las alteraciones histológicas por HE y la detección de antígeno viral por IHQ, observamos que intensa marcación celular con antígenos del VILH se observó en los dos días que antecedieron a la muerte de los animales; sin embargo el título viral fue bajo en los tejidos. Una hipótesis para explicar este hecho es que esta marcación de antígenos represente inmunocomplejos. Si realmente lo representa, se justifican los bajos títulos virales encontrados; esto contrasta, no obstante con la extensión de la lesión y la marcación celular de antígenos observadas en el cerebro.
Considerando que la lesión cerebral fue la causa de la muerte de los hámsteres en el ensayo ¿cómo explicar los bajos títulos virales encontrados? Entendemos que el virus inoculado intraperitonialmente demoró a alcanzar y establecerse en el SNC y, cuando lo hizo y empezó a replicar, el título viral observado fue bajo, pues hubo tiempo para la producción y la circulación de los anticuerpos, que, alcanzando el SNC, se INEN a los antígenos virales, lo que disminuye el título viral en el cerebro, como observado en este estudio. Es posible, sin embargo, que otros factores hayan interferido, culminando en la muerte de los animales al séptimo día p.i. Uno de ellos puede haber sido la elevada dosis del inóculo. De hecho, el inóculo de 9,8 DL50/0,02 mL es elevadísimo, lo que culminó con la muerte de todos los animales con siete días p.i. Efectivamente, en el trabajo de Tesh y colaboradores21, los autores usaron la dosis de 3,0 DL50/0,02 mL del VFA. Aun considerando que el título DL50/0,02 mL del VILH para hámsteres fue ligeramente superior, la dosis en la VIP se demostró exagerada.
El estudio sobre la patogenicidad del VILH en humanos, realizado por Southam y Moore18, demostró que la inoculación subcutánea del VILH en humanos resulta en enfermedad febril por varios días, sin embargo, en algunos casos, los pacientes desarrollaron señales de encefalitis blanda durante el curso de la enfermedad, reforzando el neurotropismo del VILH previamente documentado y confirmado en este estudio.
CONCLUSIONES
Con base en los resultados obtenidos, podemos concluir que el cerebro fue el órgano que presentó más riqueza de alteraciones teciduales, siendo que la presencia del virus en este órgano sucede en dos etapas (evolución bifásica): la primera, 24 horas luego de la infección experimental y, la segunda al quinto Con base en los resultados obtenidos, podemos concluir que el cerebro fue el órgano que presentó más riqueza de alteraciones teciduales, siendo que la presencia del virus en este órgano sucede en dos etapas (evolución bifásica): la primera, 24 horas luego de la infección experimental y, la segunda al (DL50) encontrados en esos órganos, durante la titulación en ratones, son consecuencia de la presencia del VILH en la corriente sanguínea, o sea, de la viremia; las lesiones histopatológicas graves y la presencia de gran cantidad de antígenos observada en los cerebros por IHQ, corroboradas por el título viral en este órgano, sugiere que la muerte de los animales ocurrió por encefalitis, sugiriendo que, independiente de la vía de inoculación, el VILH presenta intenso neurotropismo.
AGRADECIMIENTOS
Los autores agradecen a Basílio da Silva Buna, Luiz Roberto Oliveira da Costa, Ruth Freitas y Valter Miranda Campos por el apoyo técnico dispensado durante el desarrollo de nuestro estudio experimental.
REFERENCIAS
1 Barros VLRS. Arbovírus Morumbi (Phlebovirus: Bunyaviridae) - Estudo histopatológico e imuno-histoquímico do fígado na infecção experimental em camundongos: comparação entre as vias cerebral, intraperitoneal e subcutânea [dissertação]. Belém: Universidade Federal do Pará; 2000. [ Links ]
2 Causey OR, Causey CE, Maroja OM, Macedo DG. The isolation of arthropod-borne viruses, including members of two hitherto undescribed serological groups, in the Amazon region of Brazil. Am J Trop Med Hyg. 1961 Mar;10:227-49.
3 Clarke DH, Casals J. Techniques for hemagglutination and hemagglutination-inhibition with arthropodborne viruses. Am J Trop Med Hyg. 1958 Sep;7(5):561-73.
4 Dégallier N, Travassos da Rosa AP, Hervé JP, Vasconcelos PFC, Travassos da Rosa JF, Sá Filho GC, et al. Modifications of arbovirus eco-epidemiology in Tucuruí, Pará, Brazilian Amazônia, Related to the Construction of a Hydroeletric dam. In: Uren MF, Blok J, Manderson LH, editores. Arbovirus research in Austrália. Brisbane; 1992. p. 393.
5 Fulton F, Dumbell KR. The serological comparison of strains of Influenza Virus. J Gen Microbiol. 1949 Jan;3(1):97-111.
6 Hervé JP, Dégallier N, Travassos da Rosa AP, Pinheiro FP, Sá Filho GC. Aspectos ecológicos. In: Fundação Serviços de Saúde Pública. Instituto Evandro Chagas: 50 anos de contribuição às ciências biológicas e à medicina tropical. Belém; 1986. Vol. 1, p. 409-437. [ Links ]
7 International Committee on Taxonomy of Viruses. Management – 2006: 00.026 - Flaviviridae. In: Büchen-Osmond C, editor. The Universal Virus Database. Columbia University, New York, USA. Virus Taxonomy 2008. [Internet]. 2009 Jul [citado em 2009 jul 24]. Disponível em: http://www.ncbi.nlm.nih.gov/ICTVdb/Ictv/fs_flavi.htm. [ Links ]
8 Karabatsos N. International catalogue of arboviruses including certain other viruses of vertebrates. 3rd ed. San Antonio, USA: ASTMH; 1985. 1141p.
9 Koprowski H, Hughes TP. Physical Properties, Pathogenicity and Cultivation. J Immunol. 1946;54(4):371-385.
10 Medeiros DBA, Nunes MRT, Vasconcelos PFC, Chang GJJ, Kuno G. Complete genome characterization of Rocio virus (Flavivirus: Flaviviridae), a Brazilian flavivirus isolated from a fatal case of encephalitis during an epidemic in São Paulo State. J Gen Virol. 2007 Aug;88(8):2237-46. DOI 10.1099/vir.0.82883-0 [ Links ]
11 Pereira LE, Suzuki A, Coimbra TLM, Souza RP, Chamelet ELB. Ilheus arbovirus in wild birds (Sporophila caerulescens and Molothrus bonariensis). Rev Saude Publica. 2001 Apr;35(2):119-23.
12 Pinheiro FP, Travassos da Rosa APA. Arboviral zoonoses of Central and South America. In: Beran GW, editor. Handbook of zoonoses. 2nd ed. Boca Ranton: CRC Press; 1994. Seccion B Viral; p. 201-225.
13 Pinheiro FP, Travassos da Rosa APA, Freitas RB, Travassos da Rosa JFS, Vasconcelos PFC. Aspectos clínico-epidemiológicos das arboviroses. In: Fundação Serviços de Saúde Pública. Instituto Evandro Chagas: 50 anos de contribuição às ciências biológicas e à medicina tropical. Belém; 1986. Vol. 1, p. 375-408.
14 Prías-Landinez E, Bernal-Cubides C, Morales-Alarcón A. Isolation of Ilheus Virus from Man in Colombia. Am J Trop Med Hyg. 1968 Jan;17(1):112-4. [ Links ]
15 Prophet EB, Mills B, Arrington JB, Sobin LH. Laboratory methods in histotechnology. Washington: American registry of athology;1992. p. 3-80.
16 Reed LJ, Muench H. A simple method of estimating fifty per cent end-points. Am J Hyg. 1938 May;27(3):493-7.
17 Shope RE, Sather GE. Arbovirus. In: Lenette ED, Schmidt MJ. Diagnostic procedures for virus, ricketsias and chlamydial infections. Washington: American Public Health Association; 1979. p. 767-814.
18 Southam CM, Moore AE. West Nile, Ilheus and Bunyawera Virus Infectious in man. Am J Trop Med. 1951;31(6):724-41.
19 Spence LP, Anderson CR, Downs WG. Isolation of Ilheus virus from human beings in Trinidad, West Indies. Trans Roy Soc Trop Med Hyg. 1962;56(6):504-9. [ Links ]
20 Srihongse S, Johnson CM. The Isolation of Ilheus Virus from Man in Panamá. Am J Trop Med Hyg. 1967 Jul;16(4):516-8.
21 Tesh RB, Guzman H, Travassos da Rosa APA, Vasconcelos PFC, Dias LB, Bunnell JE, et al. Experimental yellow fever virus infection in the Golden Hamster (Mesocricetus auratus) 1. Virologic, biochemical and immunologic studies. J Infect Dis. 2001 May;183(10):1431-6. [ Links ]
22 Travassos da Rosa JFS, Travassos da Rosa APA, Vasconcelos PFC, Pinheiro FP, Rodrigues SG, Travassos da Rosa ES, et al. Arboviruses isolated in the Evandro Chagas Institute, including some described for the first time in the Brazilian Amazon region their know hosts, and their pathology for man. In: Travassos da Rosa PA, Vasconcelos PFC, Travassos da Rosa JFS, editors. Overview of arbovirology in Brazil and neighbouring countries. Belém: Instituto Evandro Chagas;1998. p. 19-31. [ Links ]
23 Vasconcelos PFC, Travassos da Rosa APA, Pinheiro FP, Shope RE, Travassos da Rosa JFS, Rodrigues SG, et al. Arboviruses pathogenic for man in Brazil. In: Travassos da Rosa APA, Vasconcelos PFC, Travassos da Rosa FS, editors. Overview of arbovirology in Brazil and neighbouring countries. Belém: Instituto Evandro Chagas; 1998. p. 72-99. [ Links ]
24 Xiao SY, Guzmán H, Travassos da Rosa APA, Zhu HB, Tesh RB. Alteration of clinical outcome and histopathology of Yellow Fever Virus infection in a hamster model by previous infection with Heterologous Flaviviruses. Am J Trop Med Hyg. 2003 Jun;68(6):695-703. [ Links ]
25 Xiao SY, Zhang H, Guzman H, Tesh RB. Experimental Yelow Fever Virus infection in the Golden Hamster (Mesocricetus auratus). II. Pathology. J Infect Dis. 2001 May;183(10):1437-44. [ Links ]
 Correspondência/Correspondence/Correspondencia:
Correspondência/Correspondence/Correspondencia:
Raimunda do Socorro da Silva Azevedo
Instituto Evandro Chagas,
Seção de Arbovirologia e Febres Hemorrágicas
Rodovia BR 316, km 7, s/no,
Bairro: Levilândia
CEP: 67030-000
Ananindeua-Pará-Brasil
E-mail:raimundaazevedo@iec.pa.gov.br
Recebido em/Received/Recibido en: 29/7/2009
Aceito em/Accepted/Aceito en: 25/9/2009
