








Servicios Personalizados
Revista

Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Pan-Amazônica de Saúde
versión impresa ISSN 2176-6215versión On-line ISSN 2176-6223
Rev Pan-Amaz Saude v.1 n.1 Ananindeua mar. 2010
http://dx.doi.org/10.5123/S2176-62232010000100023
Actualización sobre la patogénesis de la rabia
Alan C. Jackson
Departments of Internal Medicine (Neurology) and of Medical Microbiology, University of Manitoba, Winnipeg, Manitoba, Canada
Dirección para correspondencia
Endereço para correspondência
Correspondence
Título original: Rabies pathogenesis update. Traducido por: Lota Moncada.
RESUMEN
La rabia humana sigue siendo un problema de salud pública. Nuestro conocimiento sobre esa enfermedad se ha construido a través de estudios en animales de experimentación. Hay muchas preguntas sin respuesta que rodean la patogénesis de la rabia, a pesar del avance observado en los últimos tiempos. Las neuronas infectadas con rabia no pueden funcionar normalmente debido a cambios degenerativos que afectan a procesos neurales, incluyendo a dendritas y axones. Las infecciones causadas por el virus natural de la rabia no pueden eliminarse del sistema nervioso central, ya que los efectores inmunológicos no pueden ser transportados a los tejidos del cerebro, debido a la permeabilidad de la barrera hematoencefálica. No existe una terapia eficaz contra la rabia humana. El estado de coma terapéutico (inducido) ha fracasado sistemáticamente. Una mejor comprensión de la patogénesis de la rabia puede aportar nuevas ideas para el desarrollo de terapias modernas contra la rabia humana.
Palabras clave: Factores de Virulencia; Rabia; Encefalitis Viral.
INTRODUCCIÓN
La rabia humana permanece siendo un importante problema de salud pública, responsable por, como mínimo, 55 mil casos fatales por año51. La rabia humana se puede prevenir de forma eficaz, pero, permanece siendo una enfermedad casi invariablemente fatal, a despecho de la existencia de conductas terapéuticas agresivas14. Nuestro conocimiento básico acerca de la patogénesis de la rabia puede dejar algunos temas importantes sin respuesta. Una mejor comprensión de la patogénesis de la rabia puede ser muy útil para conseguir avanzar en el futuro, en la terapia para la rabia humana. En esta revisión, serán discutidos algunos asuntos seleccionados sobre la patogénesis de la rabia, con énfasis en recientes descubrimientos.
REVISIÓN DE LA PATOGÉNESIS DE LA RABIA
Mucho de lo conocido sobre la patogénesis de la rabia ha sido descubierto a partir de estudios realizados en modelos animales, generalmente, roedores infectados con cepas de virus fijos de la rabia (adaptados en laboratorio). El período de incubación de la rabia en las especies humana y animal es largo y variable, normalmente durando entre 20 y 90 días; sin embargo, en los seres humanos este período puede, en raros casos, durar más de un año44. Luego de la mordida, por un animal infectado, la saliva conteniendo el virus de la rabia se inocula en los tejidos y músculos subcutáneos. Los estudios experimentales que mejor examinaron los eventos que suceden durante el período de incubación hasta el momento presente, fueron ejecutados en zorrillos rayados americanos (Mephitis mephitis), y fue utilizado un aislado canadiense de virus rábico de calle obtenido de sus glándulas salivales4. Los estudios realizados que utilizan amplificación por transcriptasa reversa - reacción en cadena de la polimerasa (RT-PCR) mostraron que, cuando los zorrillos fueron sacrificados 62 - 64 días pos inoculación, el RNA genómico viral era encontrado, frecuentemente, en el músculo inoculado (en 4 de 9 zorrillos), sin embargo, no era encontrado en los ganglios ni en la médula espinal. Estudios inmunohistoquímicos realizados anteriormente al desarrollo de la enfermedad presentaron evidencias de infección en fibras musculares extrafusales y fibrocitos ocasionales en el punto de inoculación. Podemos concluir que el virus de la rabia está presente en el local de la mordida o en local próximo durante la mayor parte del período de incubación. La infección de las fibras musculares puede ser una fase patogenética crítica para el acceso del virus al sistema nervioso periférico.
Estudios detallados acerca de los eventos iniciales de la patogénesis en modelos "naturales" de la rabia no han sido realizados en murciélagos. Los murciélagos normalmente son responsables por más mordidas superficiales que vectores terrestres. Existe una rica inervación sensorial y autonómica en la piel y en tejidos subcutáneos, que es implicada en un período de tiempo similar al de las infecciones causadas por vectores que provocan mordidas más profundas. En Norteamérica, la mayor parte de las víctimas de rabia no presenta histórico de mordida por animales, probablemente debido al hecho de no percibirlas. En parte, este hecho es relacionado al tamaño diminuto de las mordidas - como ejemplo, podemos citar a los murciélagos de pelo plateado (silver-haired bats). La variante de virus rábico de murciélago más corrientemente responsable por la rabia humana en América del Norte está asociada a los murciélagos de pelo plateado/murciélagos orientales pipistrellus37. Las mordidas del murciélago pueden ocasionar lesiones que parecen bien comunes18. Estudios experimentales con virus de murciélagos de pelo plateado indican que se replica bien en temperaturas corporales más bajas que lo normal (34o C) y que tiene mayor poder de infestación que la variante "de calle" de coyote en tipos de células presentes en la dermis, incluyendo aquí a los fibroblastos y a las células epiteliales36. Por eso, el virus de murciélagos de pelo plateado es probablemente bien adaptado para realizar una eficiente replicación local en la dermis, lo que podría explicar el suceso de esta variante.
El virus de la rabia se liga a receptores nicotínicos de acetilcolina en la junción neuromuscular . El virus se disloca para el sistema nervioso central (SNC) dentro de axones motores y axones sensoriales por medio del transporte axonal retrógrado rápido a una tasa de 12 -100 mm por día29,34,47. El virus de la rabia ha sido utilizado como marcador neuroanatómico con el fin de definir los circuitos de neuronas ligados sinápticamente en roedores y primates. Estos estudios han demostrado que el transporte axonal del virus de la rabia ocurre exclusivamente en el sentido retrógrado45,27. Cuando las neuronas son infectadas en la médula espinal, ocurre una posterior diseminación neurona a neurona dentro de los axones en el SNC por medio del transporte axonal rápido en conexiones neuroanatómicas. Muchos tipos de células neuronales son infestadas en una amplia diseminación en el SNC; la infección de células no-neuronales ocurre en frecuencia bastante menor. La infección cerebral ocasiona cambios de comportamiento, probablemente debido a la infección de neuronas en áreas límbicas, lo que facilita la transmisión por mordidas en vectores de la rabia. Hay una diseminación del virus de la rabia distante del SNC (diseminación centrífuga) por vías neuronales, particularmente involucrando al sistema nervioso parasimpático, que es responsable por la infección de las glándulas salivares, de la pele (la biopsia de piel es un examen diagnóstico útil), del corazón y de una variedad de otros órganos14,29. El virus de la rabia infeccioso es secretado en la saliva de vectores de la rabia, lo que se configura importante para la transmisión para otros reservorios.
DISFUNCIÓN NEURONAL Y MUERTE EN INFECCIONES POR EL VIRUS DE LA RABIA
La rabia natural es normalmente caracterizada por señales
neurológicas graves y resultados fatales. Sin embargo, los cambios neuropatológicos
en el SNC son relativamente leves, consistiendo en inflamación leve,
acompañada de pequeña degeneración neuronal, corroborando
el concepto de que la disfunción neuronal, más que la muerte de
la célula neuronal, desempeña un papel importante en la producción
de la enfermedad15,25. Diversos estudios experimentales sobre la infección
por el virus de la rabia han investigado potenciales anormalidades en la neurotransmisión
involucrando acetilcolina13,48,6, serotonina3,2, y ácido 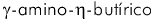 (GABA)30. Fueron encontradas anormalidades de importancia controversial, pero
ningún defecto fundamental que explicara la disfunción neuronal
en la rabia quedó demostrada.
(GABA)30. Fueron encontradas anormalidades de importancia controversial, pero
ningún defecto fundamental que explicara la disfunción neuronal
en la rabia quedó demostrada.
La disfunción de canales de iones ya fue demostrada en el virus de la rabia en cultivo de células de neuroblastoma de ratones (NA) infectadas, con el uso de la técnica de Patch-Clamp de célula entera12. La infección redujo la expresión funcional de los canales de sodio dependientes de voltaje y de los canales de potasio correctores del flujo de internalización. Hubo también una reducción en el potencial de membrana restante, reflejando su despolarización. No hubo alteración en la expresión de los canales de potasio de rectificación tardía, indicando que la disfunción no selectiva de canales iónicos no había sucedido. La reducción de los canales de sodio y de los canales de potasio correctores del flujo de internalización podría impedir neuronas infectadas de utilizar sus potenciales de acción y de generar potenciales sinápticos, lo que resultaría en perjuicio funcional.
Koprowski y colaboradores28 levantaron la hipótesis de que la neurotoxicidad del óxido nítrico podría mediar la disfunción neuronal en la rabia. La inducción de mRNA óxido nítrico sintetasa inducible28 y el aumento de los niveles de óxido nítrico en el cerebro10 ya fueron demostrados en roedores infectados por el virus de la rabia, aunque la relevancia de estos hallazgos permanece incierta. Es necesario que se estudie más el papel del óxido nítrico en la patogénesis de la rabia. La exocitoxicidad fue estudiada en cultivos primarios de neuronas infectadas por el virus de la rabia y en un modelo experimental de ratones infectados por el virus de la rabia, pero no fue constatado un papel relevante para la exocitoxicidad en la disfunción neuronal o en la muerte49.
Usando hibridación substractiva, Prosniak et al38 demostraron que la infección de ratones por el virus fijo de la rabia resulta en una disminución de la regulación de la expresión génica de cerca de 90% en comparación al cerebro normal en más de 4 veces. Cerca de 1,4% de los genes, apenas, fueron positivamente regulados, incluyendo a los genes involucrados en la regulación del metabolismo celular, en la síntesis de proteínas, en el crecimiento y la diferenciación. Usando el análisis del perfil proteómico de homogeneizados de cerebro de ratones infectados por virus de rabia "de calle" (salvaje), Dhingra et al5 percibieron que los niveles de proteínas involucrados en la homeostasis iónica estaban alterados (H+ ATPase y Na+/K+ ATPase fueron positivamente regulados, y Ca2+ ATPase fue negativamente regulado). También notaron una regulación negativa de proteínas involucradas en la fijación y fusión de vesículas sinápticas con la membrana plasmática presináptica, un proceso importante para la fisiología sináptica. Esas anormalidades pueden llevar a la disfunción neuronal.
Virus neurotrópicos pueden causar la muerte cerebral tanto por apoptosis como por necrosis9,1,7. La apoptosis depende de la síntesis de macromoléculas y requiere energía, mientras que la necrosis está asociada a una falencia energética. Cada uno de esos tipos de muerte celular está asociado a características morfológicas típicas. Se observó que la cepa viral de desafío (CVS) de virus fijo de rabia indujo a muerte celular por apoptosis en células de adenocarcinoma de próstata de ratones23, en células de neuroblastoma de ratones46, y en neuronas embrionarias del hipocampo de ratones35. La muerte de neuronas por apoptosis se observó, principalmente, en cerebros de ratones de diferentes edades, inoculados por vía intracerebral con la cepa CVS del virus rábico fijo23,46,19. Sin embargo la muerte neuronal no es prominente luego de la inoculación periférica en animales adultos43,15,39. Los virus atenuados son mejores inductores de apoptosis neuronal en cultivos de neuronas primarios, bien como en ratones luego de la inoculación periférica, que la mayor parte de las cepas virulentas35,21. Contrariamente a lo ya encontrado en anteriores estudios26, recientemente suministramos fuertes evidencias de que la apoptosis neuronal no ejerce un papel importante en lo que dice respecto a la rabia humana20. Por eso, en infecciones causadas por el virus de la rabia hay mecanismos complejos involucrados en la muerte de las células con relación a la supervivencia de neuronas, tanto in vitro como en modelos animales, usando diferentes cepas virales y vías de inoculación; no obstante, la muerte de células neuronales no se configura prominente en la rabia natural. En general, la apoptosis es un mecanismo del hospedero utilizado para limitar la diseminación viral, y no ejerce un papel fundamental en la patogénesis de la rabia15.
Durante el período prodrómico de la rabia, los pacientes pueden padecer dolor, parestesia o prurito en el lugar de la herida (generalmente ya curada), que probablemente, refleja la implicación de ganglios sensitivos locales (por ejemplo, de la raíz nerviosa espinal)14. Hay una infección neuronal, acompañada de inflamación y degeneración neuronal en los ganglios, en la rabia humana y en la mayoría de los modelos experimentales de rabia40. Esta degeneración neuronal es desproporcional a lo que se ve en el SNC del modelo murino luego de la inoculación plantar con el CVS40, y también es altamente inflamatoria, lo que sugiere un proceso inmunomediado. La degeneración neuronal no presenta ni las características morfológicas de la necrosis, ni las características morfológicas o bioquímicas de la apoptosis. En los gangliocitos ocurre inicialmente una "respuesta axónica", y, posteriormente, los gangliocitos presentan muchos compartimientos autofágicos. En estadios avanzados de degeneración, aparecen vacúolos vacíos parcialmente limitados por membrana en los gangliocitos. La autofagia es un mecanismo relevante involucrado en este proceso degenerativo neuronal.
Li y colaboradores33 sugirieron que la degeneración de procesos neuronales y la ruptura de las estructuras sinápticas pueden formar la base para la disfunción neuronal en la infección por el virus de la rabia. Ellos demostraron daños graves y la desorganización de los procesos neuronales en cortes del hipocampo de ratones teñidos con plata, en animales infectados vía intracerebral con la cepa patógena N2C del virus de la rabia. Recientemente nuestro laboratorio examinó cambios morfológicos en neuronas (con énfasis en la integridad estructural de los procesos neuronales) luego de la inoculación en la superficie plantar de la pata trasera de ratones transgénicos expresando proteína fluorescente amarilla en una subpoblación de neuronas, lo que facilita la visualización de detalles morfológicos de las dendritas, axones y terminales nerviosos presinápticos43,8. En esos ratones, la expresión de proteína fluorescente amarilla es dirigida a una subpoblación de neuronas usando el vector thy1 desprovisto de exón 3 y sus regiones "flanqueadas", y se observan fuertes señales fluorescentes en dendritas, axones y terminales nerviosas presinápticas8. En este modelo, la histopatología convencional presentó leves alteraciones inflamatorias, sin alteraciones significantes en la degeneración neuronal. No obstante, en momentos clínicos posteriores, concomitantemente al desarrollo de la enfermedad clínica neurológica, la microscopía de fluorescencia señaló anormalidades acentuadas, principalmente el surgimiento de edema que involucraban dendritas y axones de neuronas piramidales de la quinta capa cortical, una implicación severa de axones en el tronco encefálico y en el pedúnculo cerebeloso inferior y graves anormalidades que afectaban a los axones de fibras nerviosas cerebelares. El comprometimiento de las neuronas piramidales de hipocampo fue mucho menos severo, tal vez porque esas neuronas hayan sido infectadas después de la inoculación periférica en este modelo22. Los cambios estructurales pueden llevar un cierto tiempo para desarrollarse porque son mediados por una función proteica axoplásmica estructural axonal. Secciones en resina y teñidas con azul de toluidina, así como la microscopía electrónica mostraron una vacuolización en neuronas corticales que correspondían a mitocondrias emaciadas y vacuolización en los neurópilos de la corteza cerebral. Edemas axonales fueron observados. En estudio ultraestructural, también se observó vacuolización en axones y en terminales nerviosos presinápticos. Estas alteraciones morfológicas son suficientes para explicar la enfermedad clínica severa y el desenlace fatal. Ellas también suministran fuertes evidencias de que la principal deficiencia de neuronas infectadas por el virus de la rabia involucraría procesos neuronales, que no se configura aparente en análisis histopatológicas de rutina.
BARRERA HEMATOENCEFÁLICA EN LA RABIA
Hooper y colaboradores10 compararon ratones infectados con una variante letal de rabia en murciélagos de pelo plateado (SHBRV) a ratones infectados con una cepa adaptada, atenuada en laboratorio (CVS-F3) en la cual se podía observar la eliminación viral sin la existencia de secuelas. Ellos descubrieron que en ambas infecciones virales había una fuerte respuesta inmune específica al virus en la periferia del hospedero. Con todo, la invasión de células inmunes en los tejidos del SNC fue disminuida con la SHRBV debido a una incapacidad de aumento de la permeabilidad de la barrera hematoencefálica, lo que, por su vez, resultó en una falla en el clearance viral42. Ratones de un linaje SJL son menos susceptibles a la infección letal por el virus de la rabia, y presentan una mayor capacidad de mediar respuestas inflamatorias en el SNC. Las infecciones por SHBRV en estos ratones está asociada a la inducción de una mayor permeabilidad de la barrera hematoencefálica y a la inflamación del SNC, lo que resulta en una eliminación viral mayor y una tasa de supervivencia mayor41. La aplicación de hormonas esteroides en estos ratones resultó en cambios reducidos en la barrera hematoencefálica y que resultó en una mayor mortalidad41. Por eso, la permeabilidad de la barrera hematoencefálica es probablemente de gran importancia para la eliminación del virus de la rabia, y abordajes terapéuticas deben ser utilizadas para aumentar esta permeabilidad. Terapias que promuevan el encerramiento de barreras hematoencefálicas, como la administración de corticoesteroides, deben ser evitadas en el manejo de pacientes humanos acometidos de rabia.
ABORDAJES DE LA TERAPIA DE LA RABIA HUMANA
No existe una terapia establecida para pacientes humanos con rabia. Un abordaje al tratamiento de la rabia humana fue recientemente publicada24. El artículo recomendó el uso de una combinación de terapias y discutió los pros y los contras de utilizar una variedad de medicamentos específicos. En 2004, una paciente de 15 años de edad, que no había sido vacunada previamente al aparecimiento de los síntomas de la enfermedad clínica, sobrevivió a la rabia50. La joven había sido mordida en el dedo por un murciélago y no buscó atención médica o recibió cualquier tratamiento en la época. Alrededor de un mes después de mordida, la paciente desarrolló síntomas clínicos de rabia, y anticuerpos neutralizantes contra el virus de la rabia, se detectaron en su suero y líquido cefalorraquídeo. La terapia incluyó coma terapéutico (inducido) con el uso de midazolam intravenoso por siete días; se mantuvo un patrón de supresión en su electroencefalograma, y se le administró fenobarbital suplementario. También recibió terapia con ketamina y terapia antiviral, incluyendo ribavirina y amantadina. Cuando la paciente mejoró, recibió alta del hospital presentando déficit neurológico, aunque presentó una mejora neurológica posteriormente11. Este caso fue el primer registro de supervivencia a la rabia en que el individuo no había sido vacunado antes del surgimiento de los síntomas de la rabia clínica. No se sabe si la terapia con uno o más agentes específicos ejerció o no un papel importante para el resultado positivo del paciente16. Sin embargo, de aquel momento en adelante, se mínimo, 13 casos en los que los principales componentes de este abordaje (el protocolo "Milwaukee") fueron utilizados, y que resultaron en desenlaces fatales17. La inducción al coma per se no se ha demostrado útil para el tratamiento de enfermedades infecciosas en el sistema nervioso, y no hay evidencias que den soporte a este enfoque para el tratamiento de la rabia u otras encefalitis virales. Delante de lo expuesto, el coma terapéutico no se debe tornar una terapia de rutina para el tratamiento de la rabia.
El desarrollo precoz de anticuerpos neutralizantes contra el virus de la rabia en el curso clínico del paciente, lo que probablemente sucede en menos de 20% de todos los que son acometidos por rabia, probablemente contribuyó para un desenlace favorable. Los virus de la rabia de murciélago pueden ser menos neurovirulentos que las variantes caninas y otras responsables por la mayoría de los casos humanos de rabia31, y la rabia causada por las variantes caninas del virus rábico puede llevar a resultados peores que la causada por variantes de murciélago. Finalmente, la mayoría de los supervivientes presentó anticuerpos neutralizantes contra el virus de la rabia en suero y líquido cefalorraquídeo, aunque otras pruebas de laboratorio para diagnóstico, normalmente presentan un resultado negativo para el antígeno del virus de la rabia y para el RNA en líquidos y tejidos (no se hicieron pruebas a tejidos cerebrales). Eso debe suceder debido a la eficiente depuración viral.
CONCLUSIONES
Las bases de la disfunción neuronal en la rabia son complejas, pero pueden abarcar cambios degenerativos que involucren procesos neuronales, tales como dendritas y axones. Por motivos aun desconocidos, la lesión neuronal es mayor en los ganglios espinales que en las neuronas del SNC. La permeabilidad a inmunoefectores de la barrera hematoencefálica, se configura importante para la depuración viral y la recuperación del paciente con rabia. No hay ninguna terapia efectiva contra la rabia humana. Esperamos que una mejor comprensión acerca de la patogénesis de la enfermedad lleve al desarrollo de nuevas terapias para la rabia humana.
REFERENCIAS
1 Allsopp TE, Fazakerley JK. Altruistic cell suicide and the specialized case of the virus-infected nervous system. Trends Neurosci. 2000 Jul;23(7):284-90. DOI:10.1016/S0166-2236(00)01591-5 [ Links ]
2 Bouzamondo E, Ladogana A, Tsiang H. Alteration of potassium-evoked 5-HT release from virus-infected rat cortical synaptosomes. Neuro Report. 1993 May;4(5):555-8. [ Links ]
3 Ceccaldi PE, Fillion MP, Ermine A, Tsiang H, Fillion G. Rabies virus selectively alters 5-HT1 receptor subtypes in rat brain. Eur J Pharmacol. 1993 Apr;245(2):129-38. [ Links ]
4 Charlton KM, Nadin-Davis S, Casey GA, Wandeler AI. The long incubation period in rabies: delayed progression of infection in muscle at the site of exposure. Acta Neuropathol. 1997 Jul;94(1):73-7. [ Links ]
5 Dhingra V, Li X, Liu Y, Fu ZF. Proteomic profiling reveals that rabies virus infection results in differential expression of host proteins involved in ion homeostasis and synaptic physiology in the central nervous system. J Neurovirol. 2007 Apr;13(2):107-17. [ Links ]
6 Dumrongphol H, Srikiatkhachorn A, Hemachudha T, Kotchabhakdi N, Govitrapong P. Alteration of muscarinic acetylcholine receptors in rabies viral-infected dog brains. J Neurol Sci. 1996 Apr;137(1):1-6. DOI:10.1016/0022-510X(95)00275-7 [ Links ]
7 Fazakerley JK, Allsopp TE. Programmed cell death in virus infections of the nervous system. Curr Top Microbiol Immunol. 2001;253:95-119.
8 Feng G, Mellor RH, Bernstein M, Keller-Peck C, Nguyen QT, Wallace M, et al. Imaging neuronal subsets in transgenic mice expressing multiple spectral variants of GFP. Neuron. 2000 Oct;28(1):41-51. [ Links ]
9 Griffin DE, Hardwick JM. Perspective: virus infections and the death of neurons. Trends Microbiol. 1999 Apr;7(4):155-60. DOI:10.1016/S0966-842X(99)01470-5 [ Links ]
10 Hooper DC, Ohnishi ST, Kean R, Numagami Y, Dietzschold B, Koprowski H. Local nitric oxide production in viral and autoimmune diseases of the central nervous system. Proc Natl Acad Sci U S A. 1995 Jun;92(12):5312-6. [ Links ]
11 Hu WT, Willoughby RE Jr, Dhonau H, Mack KJ. Longterm follow-up after treatment of rabies by induction of coma (Letter). N Engl J Med. 2007 Aug;357(9):945-6.
12 Iwata M, Komori S, Unno T, Minamoto N, Ohashi H. Modification of membrane currents in mouse neuroblastoma cells following infection with rabies virus. Br J Pharmacol. 1999 Apr;126(8):1691-8. DOI:10.1038/sj.bjp.0702473 [ Links ]
13 Jackson AC. Cholinergic system in experimental rabies in mice. Acta Virol. 1993 Dec;37(6):502-8. [ Links ]
14 Jackson AC. Human disease. In: Jackson AC, Wunner WH, editors. Rabies. 2nd ed. London: Elsevier Academic Press; 2007. p. 309-40.
15 Jackson AC. Pathogenesis. In: Jackson AC, Wunner WH, editors. Rabies, 2nd ed. London: Elsevier Academic Press; 2007. p. 341-81.
16 Jackson AC. Recovery from rabies. N Engl J Med. 2005 Jun;352(24):2549-50.
17 Jackson AC. Update on rabies diagnosis and treatment. Curr Infect Dis Rep. 2009 Jul;11(4):296- 301. [ Links ]
18 Jackson AC, Fenton MB. Human rabies and bat bites. Lancet. 2001 May;357(9269):1714.
19 Jackson AC, Park H. Apoptotic cell death in experimental rabies in suckling mice. Acta Neuropathol. 1998 Feb;95(2):159-64. [ Links ]
20 Jackson AC, Randle E, Lawrance G, Rossiter JP. Neuronal apoptosis does not play an important role in human rabies encephalitis. J Neurovirol. 2008 Oct;14(5):368-75. DOI:10.1080/13550280802216502 [ Links ]
21 Jackson AC, Rasalingam P, Weli SC. Comparative pathogenesis of recombinant rabies vaccine strain SAD-L16 and SAD-D29 with replacement of Arg333 in the glycoprotein after peripheral inoculation of neonatal mice: less neurovirulent strain is a stronger inducer of neuronal apoptosis. Acta Neuropathol. 2006 Apr;111(4):372-8. DOI:10.1007/s00401-005-0006-z [ Links ]
22 Jackson AC, Reimer DL. Pathogenesis of experimental rabies in mice: an immunohistochemical study. Acta Neuropathol. 1989;78(2):159-65. [ Links ]
23 Jackson AC, Rossiter JP. Apoptosis plays an important role in experimental rabies virus infection. J Virol. 1997 Jul;71(7):5603-7. [ Links ]
24 Jackson AC, Warrell MJ, Rupprecht CE, Ertl HC, Dietzschold B, O'Reilly M, et al. Management of rabies in humans. Clin Infect Dis. 2003 Jan;36(1):60-3. [ Links ]
25 Jackson AC, Wunner WH. Rabies. 2nd ed. London: Elsevier Academic Press; 2007.
26 Juntrakul S, Ruangvejvorachai P, Shuangshoti S, Wacharapluesadee S, Hemachudha T. Mechanisms of escape phenomenon of spinal cord and brainstem in human rabies. BMC Infect Dis. 2005 Nov;5(1):104.
27 Kelly RM, Strick PL. Rabies as a transneuronal tracer of circuits in the central nervous system. J Neurosci Methods. 2000 Nov;103(1):63-71. DOI:10.1016/S0165-0270(00)00296-X [ Links ]
28 Koprowski H, Zheng YM, Heber-Katz E, Fraser N, Rorke L, Fu ZF, et al. In vivo expression of inducible nitric oxide synthase in experimentally induced neurologic diseases. Proc Natl Acad Sci U S A. 1993 Apr;90(7):3024-7. Erratum in: Proc Natl Acad Sci U S A. 1993 Jun;90(11):5378. [ Links ]
29 Kucera P, Dolivo M, Coulon P, Flamand A. Pathways of the early propagation of virulent and avirulent rabies strains from the eye to the brain. J Virol. 1985 Jul;55(1):158-62. [ Links ]
30 Ladogana A, Bouzamondo E, Pocchiari M, Tsiang H. Modification of tritiated γ-amino-η-butyric acid transport in rabies virus-infected primary cortical cultures. J Gen Virol. 1994 Mar;75(Pt 3):623-7. [ Links ]
31 Lafon M. Bat rabies: the Achilles heel of a viral killer? Lancet. 2005 Sep;366(9489):876-7.
32 Lentz TL, Burrage TG, Smith AL, Crick J, Tignor GH. Is the acetylcholine receptor a rabies virus receptor? Science. 1982 Jan;215(4529):182-4. [ Links ]
33 Li XQ, Sarmento L, Fu ZF. Degeneration of neuronal processes after infection with pathogenic, but not attenuated, rabies viruses. J Virol. 2005 Aug;79(15):10063-8. DOI:10.1128/JVI.79.15.10063–10068.2005 [ Links ]
34 Lycke E, Tsiang H. Rabies virus infection of cultured rat sensory neurons. J Virol. 1987 Sep;61(9):2733-41. [ Links ]
35 Morimoto K, Hooper DC, Spitsin S, Koprowski H, Dietzschold B. Pathogenicity of different rabies virus variants inversely correlates with apoptosis and rabies virus glycoprotein expression in infected primary neuron cultures. J Virol. 1999 Jan;73(1):510-8. [ Links ]
36 Morimoto K, Patel M, Corisdeo S, Hooper DC, Fu ZF, Rupprecht CE, et al. Characterization of a unique variant of bat rabies virus responsible for newly emerging human cases in North America. Proc Natl Acad Sci U S A. 1996 May;93(11):5653-8. [ Links ]
37 Noah DL, Drenzek CL, Smith JS, Krebs JW, Orciari L, Shaddock J, et al. Epidemiology of human rabies in the United States, 1980 to 1996. Ann Intern Med. 1998 Jun;128(11):922-30. [ Links ]
38 Prosniak M, Hooper DC, Dietzschold B, Koprowski H. Effect of rabies virus infection on gene expression in mouse brain. Proc Natl Acad Sci U S A. 2001 Feb;98(5):2758-63. DOI: 10.1073/pnas.051630298 [ Links ]
39 Reid JE, Jackson AC. Experimental rabies virus infection in Artibeus jamaicensis bats with CVS-24 variants. J Neurovirol. 2001 Dec;7(6):511-7. DOI: 10.1080/135502801753248097 [ Links ]
40 Rossiter JP, Hsu L, Jackson AC. Selective vulnerability of dorsal root ganglia neurons in experimental rabies after peripheral inoculation of CVS-11 in adult mice. Acta Neuropathol. 2009 Aug;118(2):249-59. [ Links ]
41 Roy A, Hooper DC. Lethal silver-haired bat rabies virus infection can be prevented by opening the blood-brain barrier. J Virol. 2007 Aug;81(15):7993-8. DOI:10.1128/JVI.00710-07 [ Links ]
42 Roy A, Phares TW, Koprowski H, Hooper DC. Failure to open the blood-brain barrier and deliver immune effectors to central nervous system tissues leads to the lethal outcome of silver-haired bat rabies virus infection. J Virol. 2007 Feb;81(3):1110-8. DOI: 10.1128/JVI.01964-06 [ Links ]
43 Scott CA, Rossiter JP, Andrew RD, Jackson AC. Structural abnormalities in neurons are sufficient to explain the clinical disease and fatal outcome of experimental rabies in yellow fluorescent protein-expressing transgenic mice. J Virol. 2008 Jan;82(1):513-21. DOI:10.1128/JVI.01677-07 [ Links ]
44 Smith JS, Fishbein DB, Rupprecht CE, Clark K. Unexplained rabies in three immigrants in the United States. A virologic investigation. N Engl J Med. 1991 Jan;324(4):205-11. [ Links ]
45 Tang Y, Rampin O, Giuliano F, Ugolini G. Spinal and brain circuits to motoneurons of the bulbospongiosus muscle: retrograde transneuronal tracing with rabies virus. J Comp Neurol. 1999 Nov;414(2):167-92. [ Links ]
46 Theerasurakarn S, Ubol S. Apoptosis induction in brain during the fixed strain of rabies virus infection correlates with onset and severity of illness. J Neurovirol. 1998 Aug;4(4):407-14. [ Links ]
47 Tsiang H, Ceccaldi PE, Lycke E. Rabies virus infection and transport in human sensory dorsal root ganglia neurons. J Gen Virol. 1991 May;72(Pt 5):1191-4.
48 Tsiang H. Neuronal function impairment in rabies-infected rat brain. J Gen Virol. 1982 Aug;61(Pt 2):277-81. [ Links ]
49 Weli SC, Scott CA, Ward CA, Jackson AC. Rabies virus infection of primary neuronal cultures and adult mice: failure to demonstrate evidence of excitotoxicity. J Virol. 2006 Oct;80(20):10270-3. DOI:10.1128/JVI.01272-06 [ Links ]
50 Willoughby RE Jr, Tieves KS, Hoffman GM, Ghanayem NS, Amlie-Lefond CM, Schwabe MJ, et al. Survival after treatment of rabies with induction of coma. N Engl J Med. 2005 Jun;352(24):2508-14. [ Links ]
51 World Health Organization. WHO expert consultation on rabies: first report. Geneva; 2005. p. 1-88.
 Correspondência/Correspondence/Correspondencia:
Correspondência/Correspondence/Correspondencia:
Alan C. Jackson
Health Sciences Centre
GF-543, 820 Sherbrook Street
Winnipeg, Manitoba R3A 1R9 Canada
Phone: 204-787-1578 Fax: 204-787-1486
E-mail: ajackson2@hsc.mb.ca
Recebido em/Received/Recibido en: 21/06/2009
Aceito em/Accepted/Aceito en: 02/10/2009