










Serviços Personalizados
Journal

Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Curriculum ScienTI
Curriculum ScienTI Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Pan-Amazônica de Saúde
versão impressa ISSN 2176-6215versão On-line ISSN 2176-6223
Rev Pan-Amaz Saude v.1 n.2 Ananindeua jun. 2010
http://dx.doi.org/10.5123/S2176-62232010000200018
Leishmania chagasi Cunha & Chagas, 1937: nativa ou introduzida? Uma breve revisão
Fernando Tobias SilveiraI; Carlos Eduardo Pereira CorbettII
ISeção de Parasitologia,
Instituto Evandro Chagas/SVS/MS, Ananindeua, Pará, Brasil. Núcleo
de Medicina Tropical, Universidade Federal do Pará, Belém,
Pará,
Brasil
IIDepartamento de Patologia, Faculdade de
Medicina da Universidade de São Paulo, São Paulo, São
Paulo, Brasil
Correspondence
Endereço para
correspondência
Dirección para correspondencia
RESUMO
Esta revisão aborda a etiologia da leishmaniose visceral americana devido a uma recente polêmica sobre a origem do seu agente etiológico, a Leishmania chagasi Cunha & Chagas, 1937. Conforme é sabido, este parasito foi descrito como uma nova espécie de Leishmania em razão da sua incapacidade de produzir, experimentalmente, a leishmaniose visceral no cão doméstico; este caráter a diferenciou de outro agente etiológico já conhecido da leishmaniose visceral na Bacia do Mediterrâneo na Europa: Leishmania infantum Nicolle, 1908. Após 50 anos da descrição da Leishmania chagasi, o gênero Leishmania sofreu ampla revisão e o parasito foi reclassificado como um membro do subgênero Leishmania, espécie Leishmania (Leishmania) chagasi. Recentemente, em seguida a uma análise molecular usando a técnica da amplificação aleatória polimórfica do DNA (RAPD), que comparou a L. (L.) chagasi com a L. (L.) infantum, foi concluído que ambos os parasitos eram geneticamente indistinguíveis e, portanto, que a L. (L.) chagasi era sinônimo de L. (L.) infantum. Por esse motivo, esta revisão procurou agregar todo o conhecimento sobre a eco-epidemiologia da L. (L.) chagasi na Amazônia brasileira, principalmente acerca dos hábitos silvestres de seu flebotomíneo vetor, Lutzomyia longipalis, e seu reservatório vertebrado, a raposa do campo Cerdocyon thous, com o propósito de demonstrar que a L. (L.) chagasi não pode ser negligenciada do cenário parasitológico da leishmaniose visceral no Novo Mundo; ela deve ser considerada, pelo menos em um nível subespecífico, como Leishmania (L.) infantum chagasi.
Palavras-chave: Leishmania; Leishmaniose Visceral; Leishmania chagasi.
REVISÃO HISTÓRICA SOBRE O STATUS TAXONÔMICO DO AGENTE ETIOLÓGICO DA LEISHMANIOSE VISCERAL AMERICANA (LVA)
O agente etiológico da leishmaniose visceral americana (LVA) foi descrito pela primeira vez por Cunha e Chagas3, tendo recebido o nome específico de Leishmania chagasi, um parasita protozoário pertencente à família Trypanosomatidae Doflein, 1901 emend Grobben, 1905 e ao gênero Leishmania Ross, 1903. A decisão de considerá-lo uma nova espécie de Leishmania deveu-se principalmente aos repetidos resultados negativos que se seguiram às tentativas de infecção experimental de cachorros com o parasita, o que o distinguiu do outro agente etiológico da leishmaniose visceral já conhecido em alguns países europeus da Bacia do Mediterrâneo, Leishmania infantum Nicolle, 1908. Entretanto, apenas cinquenta anos após a descrição de Leishmania chagasi, Lainson e Shaw10 fizeram uma profunda revisão do gênero Leishmania e reclassificaram o parasita como membro do subgénero Leishmania Saf'Janova 1982 (= Ross 1903), espécie Leishmania (Leishmania) chagasi.
Recentemente, após uma abrangente comparação entre várias cepas de L. (L.) chagasi que haviam sido isoladas de diferentes origens e países da América do Sul, particularmente do Brasil (de humanos, cachorros domésticos e da raposa selvagem Cerdocyon thous), e cepas de L. (L.) infantum originadas da área endêmica para leishmaniose visceral em países europeus da Bacia do Mediterrâneo, tais como Portugal e Espanha, a técnica de amplificação ao acaso de DNA polifórfico (RAPD) demonstrou que as sequências de DNA de ambas as espécies de parasita eram idênticas. Por esta razão, os autores daquela análise concluíram que L. (L.) chagasi é sinônimo de L. (L.) infantum e, consequentemente, propuseram que L. (L.) chagasi não deveria ser considerada uma espécie válida15.
Mais recentemente, no entanto, Lainson e Shaw11 defenderam a manutenção do parasita em um nível subespecífico, como Leishmania (L.) infantum chagasi, com base em suas características etiológicas, como o habitat silvestre de seu flebotomíneo vetor, Lutzomyia longipalpis7, e seu reservatório vertebrado natural, a raposa selvagem Cerdocyon thous20, bem como nas diferenças previamente estabelecidas entre os perfis dos fragmentos de kDNA de L. (L.) chagasi e L. (L.) infantum, demonstradas pelo uso da técnica de digestão por endonuclease de restrição6,5, de proteínas de superfície radioiodinadas de suas formas promastigotas, da geração de anticorpos monoclonais contra superfícies promastigotas18 e de estudos comparativos de radiorrespirometria4.
O ATUAL DILEMA A RESPEITO DA ORIGEM DE LEISHMANIA CHAGASI: NATIVA OU INTRODUZIDA?
A despeito dos compreensíveis argumentos apresentados por Lainson e Shaw11 em defesa da manutenção do nome científico de L. (L.) i. chagasi em um nível subespecífico, o que parece ser absolutamente justificável, tendo em vista seu longo histórico científico de mais de cinquenta anos de trabalhos publicados a respeito da ecoepidemiologia, das manifestações clínicas e da imunopatologia da doença causada por este parasita na América, alguns autores15,16,14 têm tentado desqualificar quaisquer esforços visando à manutenção deste parasita, seja em nível específico, [L. (L.) chagasi Lainson e Shaw, 1987], seja em nível subespecífico [L. (L.) i. chagasi Lainson e Shaw, 2005]. Para esses autores, apenas L. (L.) infantum e L. (L.) donovani seriam reconhecidos como agentes etiológicos da leishmaniose visceral, sendo que o primeiro ocorre nas áreas endêmicas da Bacia do Mediterrâneo, na Europa, no norte da África e nas Américas Central e do Sul, e o segundo, nas áreas endêmicas da India, leste da África e Oriente-Médio.
Diante do exposto, achamos oportuno expressar nossa opinião a respeito da proposta taxonómica de Mauricio et al15,16, que utilizou a análise molecular do DNA do parasita como o único e definitivo critério para decidir acerca da originalidade da espécie de Leishmania sob estudo. Deve-se ressaltar que a definição de uma espécie não deve ser baseada somente em suas características filogenéticas, mas também em seus aspectos biológicos, fenéticos e fenotípicos, resultantes de sua carga parasito-genómica, e suas interações com o meio ambiente. Consequentemente, concepções e tecnologias da genética moderna devem ser levadas em consideração para que estes conceitos sejam manipulados24. A este respeito, parece claro que Mauricio et al15,16 ignoraram aspectos fenotípicos tipicamente utilizados para classificar os parasitas de Leishmania, tais como: a morfologia das formas amastigotas e promastigotas; o comportamento biológico experimental do parasita em animais domésticos e selvagens, bem como em vetores flebotomíneos; e a imunologia, bioquímica, ecoepidemiologia e patogenia, ou seja, as manifestações clínicas da infecção10,19.
Em contraposição ao defendido por esses autores15,16,14, de que L. (L.) chagasi (sinónimo de L. (L.) infantum) fora introduzida na América durante a sua colonização, dois questionamentos devem ser considerados antes de qualquer afirmação. Primeiro, considerando que os primeiros casos de LVA no Brasil foram diagnosticados em 193417, como explicar que, em um curto intervalo de tempo, pouco mais de 400 anos de colonização, L. (L.) infantum tornou-se rapidamente capaz de sobreviver em Cerdocyon thous, que, diferentemente do cachorro doméstico, não apresenta quaisquer sinais ou sintomas devido à ação patogênica induzida pelo parasita? Esta adaptação reflete um estágio de coexistência entre o parasita e seu hospedeiro vertebrado geralmente aceito após um longo processo interativo entre aquele e a resposta imune deste, o que ocorre somente depois de milhares de anos. Segundo, L. (L.) infantum, que estava adaptado ao seu vetor flebotomíneo, Phlebotomus dubosqi, em países endêmicos da Europa, precisaria ter se adaptado rapidamente a outra espécie de vetor flebotomíneo, Lutzomyia longipalpis, original de um continente que apresenta condições climáticas e ecologia inteiramente diferentes das observadas no continente europeu. Ademais, é importante destacar evidências recentes sugerindo que este processo de especialização entre o parasita e seu vetor flebotomíneo é fortemente influenciado por uma interação específica entre as moléculas de glicoconjugados ligantes, especialmente o lipofosfoglicano (LPG), presentes na membrana plasmática de superfície das formas promastigotas metacíclicas das espécies de Leishmania e seus receptores na membrana das células epiteliais da parede do intestino médio do vetor flebotomíneo2,22,21. O LPG foi implicado como uma molécula de adesão específica que intermedeia a interação entre Leishmania e o epitélio do intestino médio do vetor flebotomíneo. Diante disto, dever-se-ia supor que L. (L.) infantum, que é transmitido naturalmente por Phlebotomus dubosqi, seria capaz de se adaptar rapidamente ao epitélio do intestino médio de Lutzomyia longipalpis, um vetor flebotomíneo de espécie e gênero completamente diferentes de L. (L.) infantum encontrado em uma área endêmica na Europa.
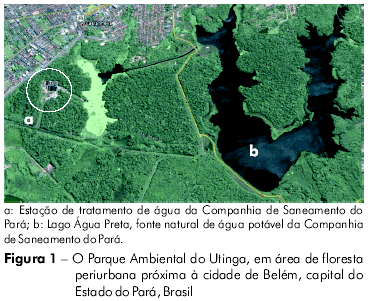
Como afirmado anteriormente, este processo de especialização entre o parasita, seu vetor biológico e seu reservatório selvagem, que caracteriza um ciclo enzoótico natural das espécies de Leishmania, foi bastante fundamentado no caso de um parasita de Leishmania ser muito próximo de um parasita de L. (L.) chagasi isolado das vísceras (fígado e baço) de três raposas caranguejeiras selvagens Cerdocyon thous saudáveis, capturadas em uma área de floresta periurbana denominada "Parque Ambiental do Utinga" (Figura 1), próximo a Belém, Estado do Pará, Amazônia Brasileira, onde não há evidências de doenças em humanos ou em caninos até o presente momento. No entanto, já foi comprovado que este parasita de Leishmania causa infecção visceral letal em hamsters dourados após três meses de inoculação via intraperitonial12. Portanto, considerando que esta área ainda não é habitada nem por humanos nem por cachorros, esta descoberta representa uma forte evidência da existência de um ciclo enzoótico indígena de parasitas de L. (L.) chagasi-like na Amazônia Brasileira, que certamente teve origem antes do histórico recente da colonização do continente americano.
Outro ponto que contradiz a hipótese evolucionária15,16,14, que concorda com a tese de que o gênero Leishmania teve origem na América do Sul no Paleoceno ou Eoceno, com posterior diversificação após sua migração para a Ásia, diz respeito à interpretação do que ocorreu com o ancestral de Leishmania que migrou para o continente asiático. A hipótese evolucionária afirma que nenhum ancestral de Leishmania permaneceu na América do Sul. No entanto, como explicar a origem de um vasto número de espécies de Leishmania que existem na região? Na verdade, há mais de vinte espécies de Leishmania reconhecidas pertencentes aos subgêneros Leishmania e Viannia, incluindo L. (L.) chagasi, o único parasita de Leishmania responsável pela LVA11.
CONSIDERAÇÕES FINAIS
Considerando os argumentos acima mencionados, dois aspectos importantes devem ser destacados. O primeiro refere-se à considerável especulação acerca da hipótese evolucionária proposta por Lukes et al14, baseada na análise filogenética entre L. (L.) chagasi e L. (L.) infantum. Como já dissemos, apesar de ter sido demonstrada uma grande semelhança filogenética entre estes dois parasitas, os autores propuseram que o gênero Leishmania teve origem na América do Sul, no Paleoceno ou Eoceno, com posterior diversificação após sua migração para a Ásia. Este fato produziu duas ramificações: Leishmania donovani, no subcontinente indiano, e Leishmania infantum, no continente europeu. Mais recentemente, foi também sugerida que Leishmania infantum teria sido introduzido na América por seus colonizadores. Esta hipótese evolucionária parece ser especulativa e contraditória: é impossível acreditar que um ancestral de Leishmania migraria para o continente asiático sem deixar um parasita descendente em seu lugar de origem. Recentemente, no Parque Ambiental do Utinga, na mesma área em que Lainson et al12 encontraram um parasita L. (L.) chagasi-like de uma infecção visceral natural em uma raposa Cerdocyon thous, um parasita de Leishmania foi identificado em Lutzomyia tuberculata, Leishmania (Viannia) utingensis Braga, Lainson, Ishikawa e Shaw1. Não há registros deste parasita fora da localidade tipo em que fora isolado pela primeira vez, o que demonstra claramente que, como em outros parasitas de L. (L.) chagasi-like, existem ciclos enzoóticos indígenas fechados de parasitas de Leishmania no Novo Mundo que indubitavelmente descenderam diretamente de um ancestral indígena de Leishmania. De acordo com estes dados, também deve ser também destacada a descoberta recente de Lutzomyia longipalpis em uma área de floresta dentro da sede do Instituto Evandro Chagas, no Município de Ananindeua, Estado do Pará, Brasil23, área esta contígua ao Parque Ambiental do Utinga. Este fato confirma a existência de um ciclo enzoótico indígena de L. (L.) chagasi nesta área, em que não há qualquer evidência de doença visceral humana ou canina.
O segundo aspecto a ser considerado são os critérios (morfologia, biologia experimental, ecoepidemiologia, imunologia, bioquímica e patogenicidade) utilizados para a identificação e classificação dos parasitas de Leishmania, amplamente discutidos por Lainson e Shaw10 em sua última revisão do gênero Leishmania. Como recentemente observado por Tibayrenc24, apesar de terem sido reconhecidas grandes vantagens na análise do DNA de Leishmania por meio de técnicas de biologia molecular para diferentes objetivos, a análise de DNA deve ser utilizada como uma ferramenta complementar para a identificação precisa de uma espécie de Leishmania, e não como um fator determinante em sua identificação. Deste modo, não podemos omitir a importância de alguns estudos acerca, principalmente, da originalidade da biologia e ecoepidemiologia de L. (L.) chagasi na América7,20,12,13,9,8.
Em conclusão, não concordamos com a proposta de desqualificar L. (L.) chagasi, que tem sido reconhecido por mais de cinquenta anos como o agente etiológico de LVA, com base na similaridade de seu DNA com o de L. (L.) infantum. Esta descoberta, sozinha, não confirma suas características taxonómicas de forma suficiente para anular todo o conhecimento científico acumulado sobre este parasita de Leishmania nos últimos cinquenta anos. Portanto, cremos que a melhor decisão a ser tomada a respeito deste assunto seria a de manter o status subespecífico do agente etiológico de LVA como L. (L.) i. chagasi, como proposto por Lainson e Shaw11 em sua última revisão dos parasitas neotropicais de Leishmania com importância médica.
REFERÊNCIAS
1 Braga RR, Lainson R, Ishikawa EA, Shaw JJ. Leishmania (Viannia) utingensis n. sp., a parasite from the sandfly Lutzomyia (Viannamyia) tuberculata in Amazonian Brazil. Parasite. 2003 Jun;10(2):111-8. [ Links ]
2 Butcher BA, Turco SJ, Hilty BA, Pimenta PF, Panunzio M, Sacks DL. Deficiency in β1,3-galactosyltransferase of a Leishmania major lipophosphoglycan mutant adversely influences the Leishmania-sand fly interaction. J Biol Chem. 1996 Aug;271(34):20573-9. [ Links ]
3 Cunha AM, Chagas E. Nova espécie de protozoário do gênero Leishmania patogênico para o homem. Leishmania chagasi n.sp. Nota prévia. Hospital (Rio J) 1937;11:3-9.
4 Decker-Jackson FE, Tang DB. Identification of Leishmania spp. by radiorespirometry: II. A statistical method of data analysis to evaluate the reproductibility and sensitivity of the technique. In: Chance ML, Walton BC, editors. Biochemical Characterization of Leishmania. Geneva: Pan American Health Organization; 1982. p. 205-45.
5 Jackson PR, Stiteler JM, Wohlieter JA, Reed SG, Badaró R, Inverso JA, et al. Characterization of Leishmania responsible for visceral disease in Brazil by restriction endonuclease digestion and hybridization of kinetoplastic DNA. In: 11th Congress International Tropical Medicine and Malaria, Calgary, Canada; 1984. p. 68.
6 Jackson PR, Wohlieter JA, Hockmeyer WT. Leishmania characterization by restriction endonuclease digestion of kinetoplastic DNA. 5th Congress International Parasitology, Toronto, Canada; 1982. p. 342.
7 Lainson R, Dye C, Shaw JJ, MacDonald DW, Courtenay O, Souza AAA, et al. Amazonian visceral leishmaniasis - distribution of the vector Lutzomyia longipalpis (Lutz & Neiva) in relation to the fox Cerdocyon thous (Linn.) and the efficiency of this reservoir host as a source of infection. Mem Inst Oswaldo Cruz. 1990 Jan-Mar;85(1):135-7. Doi:10.1590/S0074-02761990000100027 [ Links ]
8 Lainson R, Ishikawa EA, Silveira FT. American visceral leishmaniasis: wild animal hosts. Trans Roy Soc Trop Med Hyg. 2002 Nov-Dec;96:(6)30-1. [ Links ]
9 Lainson R, Shaw JJ. A brief history of the genus Leishmania (Protozoa: Kinetoplastida) in the Americas with particular reference to Amazonian Brazil. Cienc Cult. 1992 Mar-Abr;44(2-3):94-106. [ Links ]
10 Lainson R, Shaw JJ. Evolution, classification and geographical distribution. In: Peteers W, Killick-Kendrick R, editors. The Leishmaniasis in Biology and Medicine. London: Press Inc; 1987. p. 12-120.
11 Lainson R, Shaw JJ. Leishmaniasis in the new world. In: Collier L, Balows A, Sussman M, editors. Topley & Wilson's Microbiology and Microbial Infections, 10th ed. London: E Arnold; 2005. p. 313-49.
12 Lainson R, Shaw JJ, Lins ZC. Leishmaniasis in Brazil: IV The fox, Cerdocyon thous (L) as a reservoir of Leishmania donovani in Para State, Brazil. Trans R Soc Trop Med Hyg. 1969;63(6):741-5. [ Links ]
13 Lainson R, Shaw JJ, Silveira FT, Braga RR. American visceral leishmaniasis: on the origin of Leishmania (Leishmania) chagasi. Trans R Soc Trop Med Hyg. 1987; 81(3):517.
14 Lukes J, Maurício IL, Schönian G, Dujardin JC, Soteriadou K, Dedet JP, et al. Evolutionary and geographical history of the Leishmania donovani complex with a revision of current taxonomy. Proc Natl Acad Sci U S A. 2007 May;104(22):9375-80. [ Links ]
15 Mauricio IL, Howard MK, Stothard JR, Miles MA. Genomic diversity in the Leishmania donovani complex. Parasitology. 1999 Sep;119(Pt 3):237-46. Doi:10.1017/S0031182099004710 [ Links ]
16 Mauricio IL, Stothard JR, Miles MA. The strange case of Leishmania chagasi. Parasitol Today. 2000 May;16(5):188-9.
17 Penna HA. Leishmaniose visceral no Brasil. Bras Med. 1934;48:949-50.
18 Santoro F, Lemesre JL, Rizvi FS, Afchain D, Sadogirsky M. Spécificité au niveau de proteins de surface des promastigotas de Leishmania donovani (Laveran et Mesnil, 1903), Leishmania infantum Nicolle, 1908 et Leishmania chagasi Cunha et Chagas, 1937. In: Rioux JA, editor. Leishmania. Taxonomie et Phylogenese. Applications Éco-Épidémiologiques. Colloque International; 1984 July 2-6; Montpellier: L'Institut Méditerranéen d'Études Épidémiologiques et Écologiques; 1986. p. 71-6.
19 Silveira FT, Ishikawa EA, Souza AAA, Lainson R. An outbreak of cutaneous leishmaniasis among soldiers in Belém, Pará State, Brazil, caused by Leishmania (Viannia) lindenbergi n. sp. A new leishmanial parasite of man in the Amazon region. Parasite. 2002 Mar;9(1):43-50. [ Links ]
20 Silveira FT, Lainson R, Shaw JJ, Póvoa MM. Leishmaniasis in Brazil: XVIII. Further evidence incriminating the fox Cerdocyon thous (L) as a reservoir of Amazonian visceral leshmaniasis. Trans Roy Soc Trop Med Hyg. 1982;76(6):830-2.
21 Soares RP, Cardoso TL, Barron T, Araã MS, Pimente PF, Turco SJ. Leishmania braziliensis: a novel mechanism in the lipophosphoglycan regulation during metacyclogenesis. Int J Parasitol. 2005 Mar;35(3):245-53. Doi:10.1016/j.ijpara.2004.12.008 [ Links ]
22 Soares RP, Macedo ME, Ropert C, Gontijo NF, Almeida IC, Gazzinelli RT. Leishmania chagasi: lipophosphoglycan characterization and binding to the midgut of the sand fly vector Lutzomyia longipalpis. Mol Biochem Parasitol. 2002 May;121(2):213-24. [ Links ]
23 Souza AAA, Silveira FT, Silva FMM, Silva LA, Leão EF, Barata IR, et al. Flebotomíneos capturados e identificados no Campus II do Instituto Evandro Chagas - Ananindeua - Estado do Pará, Brasil. Rev Soc Bras Med Trop. 2009;42 Supl 1:441.
24 Tibayrenc M. The species concept in parasites and other pathogens: a pragmatic approach? Trends in Parasitol. 2006 Feb;22(2):66-70.Doi:10.1016/j.pt.2005.12.010 [ Links ]
 Correspondência
/ Correspondence / Correspondencia:
Correspondência
/ Correspondence / Correspondencia:
Fernando Tobias Silveira
Instituto Evandro Chagas,
Seção de Parasitologia
Rodovia BR 316, km 07, s/no. Bairro: Levilândia
CEP: 67030-000
Ananindeua-Pará-Brasil
E-mail:fernandotobias@iec.pa.gov.br
Recebido em / Received / Recibido en: 21/1/2010
Aceito em / Accepted / Aceito en: 25/6/2010
