








Servicios Personalizados
Revista

Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Curriculum ScienTI
Curriculum ScienTI Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Pan-Amazônica de Saúde
versión impresa ISSN 2176-6215versión On-line ISSN 2176-6223
Rev Pan-Amaz Saude v.1 n.4 Ananindeua dic. 2010
http://dx.doi.org/10.5123/S2176-62232010000400008
ORIGINAL ARTICLE | ARTIGO ORIGINAL | ARTÍCULO ORIGINAL
The frequency of HLA-DRB1 polymorphisms in Brazilian Plasmodium vivax malaria patients and in blood donors from the Amazon Region
Frequência de polimorfismos HLA-DRB1 em pacientes brasileiros com malária por Plasmodium vivaxe em doadores de sangue da Região Amazônica
Frecuencia de polimorfismos HLA-DRB1 en pacientes brasileños con malaria por Plasmodium vivax y en donantes de sangre de la Región Amazónica
Luciane Moreno Storti-MeloI; Daniela Reis da CostaI; Wanessa Christina Souza-NeirasI; Gustavo Capatti CassianoI; Cláudia Regina Bonini-DomingosI; Andrea Regina Baptista RossitII; Luiz Carlos de MattosIII; Ricardo Luiz Dantas MachadoIV
IUniversidade Estadual Paulista "Júlio de Mesquita Filho", São José do Rio Preto, São Paulo, Brasil
IIInstituto Biomédico, Universidade Federal Fluminense, Niterói, Rio de Janeiro, Brasil
IIILaboratório de Imunohematologia, Faculdade de Medicina de São José do Rio Preto, São José do Rio Preto, São Paulo, Brasil
IVCentro de Investigação de Microrganismos, Faculdade de Medicina de São José do Rio Preto, São José do Rio Preto, São Paulo, Brasil
Endereço para correspondência
Correspondence
Dirección para correspondencia
ABSTRACT
We evaluated the frequency of different HLA-DRB1 alleles in Plasmodium vivax-infected individuals and in healthy blood donors from malaria endemic areas of Brazil. Low-resolution human leukocyte antigen-DRB1 genotyping was performed for 73 malaria patients and 29 healthy blood donors. The most frequent alleles in individuals from northern Brazil were human leukocyte antigen-DRB1*04, *08, *07 and *13. The frequency of human leukocyte antigen-DRB1*07 was higher in malaria-infected individuals than in the control group, which reinforces the theory that this allele plays an important role in susceptibility to malaria. This study offers new information about a potential susceptibility factor for P. vivax malaria in a Brazilian population that is naturally exposed to malaria.
Keywords: Malaria; Plasmodium vivax; MHC Class II genes.
RESUMO
Este estudo avaliou a frequência de diferentes alelos HLA-DRB1 em indivíduos infectados por Plasmodium vivax e em doadores de sangue saudáveis provenientes de áreas endêmicas de malária do Brasil. Foi realizada uma genotipagem de baixa resolução dos alelos HLA-DRB1 em 73 pacientes com malária e em 29 doadores de sangue saudáveis. Os alelos mais frequentes em indivíduos do norte do Brasil foram HLA-DRB1 *04, *08, *07 e *13. A frequência de HLA-DRB1 *07 foi maior nos indivíduos infectados com malária do que no grupo controle, o que reforça a hipótese de que esse alelo desempenha um papel importante na suscetibilidade à malária. Esta pesquisa fornece novas informações sobre um fator potencial de suscetibilidade à malária por P. vivax em uma população brasileira naturalmente exposta à doença.
Palavras-chave: Malária; Plasmodium vivax; Genes Classe II do Complexo de Histocompatibilidade (MHC).
RESUMEN
Este estudio evaluó la frecuencia de diferentes alelos HLA-DRB1 en individuos infectados por Plasmodium vivax y en donantes de sangre saludables provenientes de áreas endémicas de malaria de Brasil. Se realizó un genotipado de baja resolución de los alelos HLA-DRB1 en 73 pacientes con malaria y en 29 donantes de sangre saludables. Los alelos más frecuentes en individuos del norte de Brasil fueron HLA-DRB1*04, *08, *07 y *13. La frecuencia de HLA-DRB1*07 fue más grande en individuos infectados con malaria que en el grupo control, lo que refuerza la hipótesis de que ese alelo desempeña un papel importante en la susceptibilidad a la malaria. Esta investigación suministra nuevas informaciones sobre un factor potencial de susceptibilidad a la malaria por P. vivax en una población brasileña naturalmente expuesta a la enfermedad.
Palabras clave: Malaria; Plasmodium vivax; Genes Clase II del Complejo de Histocompatibilidad (MHC).
INTRODUCTION
Human leukocyte antigen (HLA) class II genes were originally described as genes involved in immune response because different alleles of these genes are known to influence antibody production1. Studies have associated these alleles with diseases of the immune system, such as leukemia2 and vitiligo3. Moreover, population studies have reported the association of certain HLA alleles with susceptibility or resistance to infectious diseases, including leprosy4, mucosal leishmaniasis5, tuberculosis6 and hepatosplenomegaly in schistosomiasis7. Few studies have examined the influence of HLA alleles on immunity to malaria. The authors of two studies (published in 1989) were unable to establish a relationship between the HLA-DR types and the immune response to circumsporozoite protein (CSP) of Plasmodium falciparum8 and to sporozoite and gametocytes epitopes in Papua New Guinea9. However, two years later, protection against severe P. falciparum malaria was found to be associated with the presence of the HLA-Bw53, DRB1*1302 and DQB1*0501 alleles in African children10. By the turn of the 21st century, some HLA-DR alleles had been associated with an increased antibody response to Nt47 (p126 amino-terminal portion)11, to apical membrane antigen-1 (AMA-1)12 of P. falciparum and to the VK247 CSP repetition of P. vivax13.
The majority of studies investigating the influence of HLA alleles on the immune response to malaria have been conducted with P. falciparum antigens because of the higher mortality attributed to malaria caused by this parasite. However, P. vivax is the species responsible for the majority of malaria cases in South and Central America. In Brazil, this species accounts for more than 80% of the clinical cases reported in the Amazon Region14. Thus, it is important to understand the factors that control the immune response to vaccine candidate antigens, such as the asexual-blood stage proteins of P. vivax, in Brazilian people naturally exposed to the parasite. In this study, we evaluated the HLA-DRB1 allelic frequency in samples from individuals infected with P. vivax and from healthy blood donors in malaria endemic areas of Brazil.
MATERIALS AND METHODS
STUDY SUBJECTS AND LOCATIONS
The patients who were enrolled in this study met the following criteria: they sought medical assistance for clinical malaria symptoms, were over 18 years old and had a positive P. vivax malaria diagnosis by thick blood film. Signed, written informed consent was obtained before peripheral blood samples were collected from each patient. The study subjects were distributed among four Brazilian malaria endemic areas: Macapá, Amapá State (00o02'20" S, 51o03'59" W); Novo Repartimento, Pará State (04o19'50" S, 49o47'47" W); Porto Velho, Rondônia State (-08o45'43" S, 63o54'14" W); and Plácido de Castro, Acre State (10o16'33" S, 67o09'00" W). Macapá is the capital of the State of Amapá, which is located on the banks of the Amazon River, in a tropical forest region. The estimated population of Macapá is 366,486 inhabitants, and its annual parasitic index (API) was 6.0 in 2009. Porto Velho is in the State of Rondônia, located in the upper Amazon River Basin. In 2010, Porto Velho had approximately 383,425 inhabitants and an API of 53.7. Plácido de Castro is located at the border of the States of Rondônia and Amazonas, and it has a population of 18,235 inhabitants. The API of Plácido de Castro was 20.6 in 2009. Novo Repartimento is a gold mining area in the southeast of the State of Pará. Its population was approximately 55,759 inhabitants and it had an API of 15.4 in 2010. These areas are characterized by having a tropical climate without a dry season; the mean monthly precipitation level is at least 60 mm.
Subjects in the control group were blood donors who, according to the Brazilian blood bank policy and the inclusion requirements for this study, met the following criteria: they were over 18 years old; their place of birth was within the study area; they reported never having suffered from malaria attacks and had no signs of malaria during the initial interview; and they had negative results for thick blood film. Molecular diagnosis methods were used for HLA-DRB frequency comparisons. The subjects of the two groups showed no statistically significant differences in mean age or ethnicity, indicating a well-matched population (Fisher's exact test, p > 0.05). DNA samples were extracted from frozen peripheral blood, using the Easy-DNATM extraction kit (Invitrogen, Carlsbad, CA, USA); malaria diagnosis was confirmed using a semi-nested polymerase chain reaction (PCR) with specific small-subunit rDNA primers15. The protocol for this study was reviewed and approved by the Research Board of the Faculdade de Medicina de São José do Rio Preto, São Paulo State, Brazil (Process number 235/2006).
HLA CLASS II ALLELES
DNA samples from the malaria patients (n = 73) and the control group (n = 29) were subjected to genotyping of the HLA-DRB1 alleles. The DNA concentration was measured using a spectrophotometer at 260 and 280 nm, and a concentration of 100 ng/mL was used for low-resolution genotyping of HLA-DRB1 by PCR with sequence-specific primers (PCR-SSP), as previously described16.
STATISTICAL ANALYSIS
Allele frequencies were calculated with the formula AF = a/N, in which a represents the number of positive samples for a specific allele and N represents the total number of alleles in the study population17. R version 2.8.1 statistical software (The R Foundation for Statistical Computing, Vienna, Austria, http://www.r-project.org) was used for the statistical analyses. The Hardy-Weinberg equilibrium was tested according to Guo and Thompson18. The heterogeneity of HLA allelic frequencies between control and malaria-infected groups was evaluated using a Chi-square analysis, with Yate's correction test or Fisher's exact test. P-values less than 0.05 were considered statistically significant.
RESULTS
HLA-DRB1 genotyping was performed in a total of 102 individuals, including malaria-infected individuals and uninfected controls, from four endemic areas in the north of Brazil. As summarized in table 1, 13 different alleles were found; the frequencies observed for each individual and for each group are also described in table 1. The most frequent alleles for both groups were HLA-DRB1*04, *08, *07 and *13. In malaria-infected individuals, HLA-DRB1*04, *08 and *13 were the most frequent alleles, followed by HLA-DRB1*07. In the control group, HLA-DRB1*04, *08 and *13 were also the most frequent, but HLA-DRB1*07 was one of the least frequent alleles in this group. When we compared the malaria-infected group with the control group, the HLA-DRB1*07 frequency was higher in the infected group than in the control group; this difference was statistically significant (p = 0.006, Chi-square test with Yate's correction). The loci were in Hardy-Weinberg equilibrium in the studied population (Pearson's x2 = 0.5, p = 0.799).
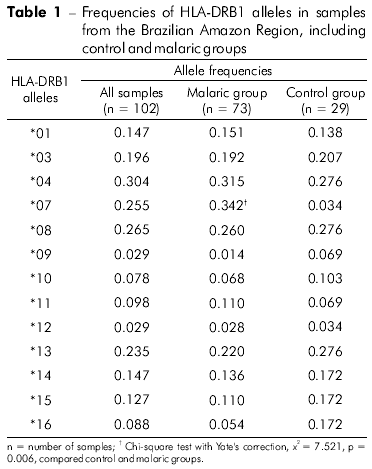
DISCUSSION
HLA class II molecules are crucial components of the adaptive immune response, and a correlation between HLA alleles and susceptibility to several diseases, including malaria, has been documented2,3,4,5,6,7,8,9,10,11,12,13. Moreover, ethnic and/or geographic variations apparently play a major role in this correlation. The nation of Brazil is as large as an entire continent, and its inhabitants are genetically diverse. This diversity results in a mixed population formed by the contributions of three important groups: Caucasians, Africans and Amerindians19. Due to the diverse history of colonization, different regions of the country have different levels of prevalence of the three ethnic groups20,21. We found a high frequency of HLA-DRB1*04 in our subjects, which is similar to the results seen in previous studies22,23. Because this allele is characteristic of indigenous Americans, its prevalence reflects the Amerindian contribution to the Brazilian population, especially in the north of the country24. Additionally, the high frequency of HLA-DRB1*04 that was observed in both malaria-infected and healthy individuals indicates that this allele is not associated with susceptibility to or protection from P. vivax malaria. On the other hand, the significantly higher frequency of the HLA-DRB1*07 in the infected group suggests a role for this allele in susceptibility to malaria. Oliveira-Ferreira et al13 detected a poor immune response induced by HLA-DRB1*07 against the VK210 repetitive region of P. vivax CSP In fact, studies with viral vaccine22,25 and vaccine trials with CSP of P. falciparum26 have shown that this allele fails to trigger a consistent immune response.
Although differences have been observed in the HLA-DRB1*07 frequencies between malaria-infected and uninfected individuals, we have not established a mechanism by which this allele may function in malaria susceptibility. There may be multiple HLA molecules associated with human malaria, although we were only able to analyze the HLA-DRB1 alleles in this study. Analysis of a single HLA molecule cannot fully reflect how multiple genetic variations impact the MHC region. One possibility is that MHC alleles could be in linkage disequilibrium with other relevant genes involved in susceptibility to P. vivax malaria. Additionally, patients homozygous for the HLA-DRB1*07 allele could not be distinguished from carriers due to the sampling limitations of this study.
In the Brazilian Amazon Region, malaria predominates in mesoendemic conditions with a wide variation in transmission14,27. Thus, malaria endemicity could be viewed as a selective pressure for maintenance of the genotype frequencies of the HLA molecules. Investigation of other malaria-associated HLA alleles, using serological, clinical and epidemiological analysis of a large sample size, is necessary. Such a study should also evaluate the influence of other infectious diseases. The epidemiology of infectious diseases is conditioned by a complex interrelationship between the parasite, its vector and the human being. The parasite-host coevolutionary process can be viewed as an arms race, in which adaptive genetic changes in one are eventually matched by alterations in the other, in this case, within the genetically diverse Amazonian populations28.
CONCLUSION
We found that the HLA-DRB1*07 allele frequency was higher in malaria patients than in the control group, which could suggest a role for this allele in susceptibility of the immune system to P. vivax malaria. Our results reinforce the need for studies that focus on genes linked to the histocompatibility complex; there is a particular need for studies using a larger sample size, as well as individuals from areas with different rates of infection, to better establish the role of the HLA in susceptibility to P. vivax parasites in this region.
ACKNOWLEDGMENTS
To all individuals enrolled in this study. We thank Drs. Carlos Eugênio Cavasini, Aline Barroso, Maria Cristina Figueredo and Mauro Tada for helping in the malaria field work. To Juliana Cintra for assistance in HLA genotyping analysis. To Dr. Antonio José Cordeiro for his participation in statistical analysis. To Professor Luiz Hildebrando Pereira da Silva for allowing the use of all facilities at Centro de Pesquisas em Medicina Tropical (Cepem).
FINANCIAL SUPPORT
This work was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo, São Paulo State, Brazil (02/09546-1; 06/00982-4) and Conselho Nacional para o Desenvolvimento Científico e Tecnológico, Brasília, Brazil (302353/03-8; 410405/2006-0).
REFERENCES
1 Germain RN. Antigen processing and presentation. In: Paul WE, editor. Fundamental Immunology. 4th ed. Philadelphia: Lippincott-Raven; 1999. p. 287-340.
2 Ozdilli K, Oguz FS, Anak S, Kekik C, Carin M, Gedikoglu G. The frequency of HLA class I and II alleles in Turkish childhood acute leukaemia patients. J Int Med Res. 2010 Sep-Oct;38(5):1835-44. [ Links ]
3 Hu DY, Ren YQ, Zhu KJ, Lv YM, Cheng H, Zhang Z, et al. Comparisons of clinical features of HLA-DRB1*07 positive and negative vitiligo patients in Chinese Han population. J Eur Acad Dermatol Venereol. In Press 2011. Doi:10.1111/j.1468-3083.2010.03971.x [ Links ]
4 Todd JR, West BC, McDonald JC. Human leukocyte antigen and leprosy: study in northern Louisiana and review. Rev Infect Dis. 1990 Jan-Fev;12(1):63-74. [ Links ]
5 Petzl-Erler ML, Belich MP, Queiroz-Telles F. Association of mucosal leishmaniasis with HLA. Hum Immunol. 1991 Dec;32(4):254-60. D![]() oi:10.1016/0198-8859(91)90088-Q [ Links ]
oi:10.1016/0198-8859(91)90088-Q [ Links ]
6 Duarte R, Carvalho C, Pereira C, Bettencourt A, Carvalho A, Villar M, et al. HLA class II alleles as markers of tuberculosis susceptibility and resistance. Rev Port Pneumol. 2011 Jan;17(1):15-9. [ Links ]
7 Assaad-Khalil SH, Helmy MA, Zaki A, Mikhail MM, el-Hai MA, el-Sawy M. Some genetic, clinical and immunologic interrelations in schistosomiasis mansoni. Ann Biol Clin (Paris). 1993;51(6):619-26. [ Links ]
8 Brown AE, Chandanayingyong D, Fuggle SV, Webster HK. Immune responses to the circumsporozoite protein of Plasmodium falciparum in relation to HLA-DR type. Tissue Antigens. 1989 Sep;34(3):200-4. [ Links ]
9 Graves PM, Bhatia K, Burkot TR, Prasad M, Wirtz RA, Beckers P. Association between HLA type and antibody response to malaria sporozoite and gametocyte epitopes is not evident in immune Papua New Guineans. Clin Exp Immunol. 1989 Dec;78(3):418- 23. [ Links ]
10 Hill AVS, Yates SNR, Allsopp CEM, Gupta S, Gilbert SC, Lalvani A, et al. Human leucocyte antigens and natural selection by malaria. Philos Trans R Soc Lond B Biol Sci. 1994 Nov;346(1317):379-85. [ Links ]
11 Banic DM, Goldberg AC, Pratt-Riccio LR, Oliveira-Ferreira J, Santos F, Gras-Masse H, et al. Human leukocyte antigen class II control of the immune response to p126-derived amino terminal peptide from Plasmodium falciparum. Am J Trop Med Hyg. 2002 May;66(5):509-15. [ Links ]
12 Johnson AH, Lee RG, Mendell NR, Shon D, Suh YJ, Bomba-Nkolo D, et al. Human leukocyte antigen class II alleles influence levels of antibodies to the Plasmodium falciparum asexual-stage apical membrane antigen 1 but not to merozoite surface antigen 2 and merozoite surface protein 1. Infect Immun. 2004 May;72(5):2762-71 . Doi:10.1128/IAI.72.5.2762-2771.2004. [ Links ]
13 Oliveira-Ferreira J, Pratt-Riccio LR, Arruda M, Santos F, Ribeiro CT, Goldberg AC, et al. HLA class II and antibody responses to circumsporozoite protein repeats of P. vivax (VK210, VK247 and P. vivax-like) in individuals naturally exposed to malaria. Acta Trop. 2004 Sep;92(1):63-9. Doi:10.1016/J.Acta tropica.2004.02.011 [ Links ]
14 Oliveira-Ferreira J, Lacerda MVG, Brasil P, Ladislau JLB, Tauil PL, Daniel-Ribeiro CT. Malaria in Brazil: an overview. Malar J. 2010;9:115. Doi:10.1186/1475-2875-9-115. [ Links ]
15 Kimura M, Kaneko O, Liu Q, Zhou M, Kawamoto F, Wataya Y, et al. Identification of the four species of human malaria parasites by nested PCR that targets variant sequences in the small subunit rRNA gene. Parasitol Intern. 1997 Jul;46(2):91-5. Doi:10.1016/s1383-5769(97)00013-5 [ Links ]
16 Olerup O, Zetterquist H. HLA-DR typing by PCR amplification with sequence-specific primers (PCR-SSP) in 2 hours: an alternative to serological DR typing in clinical practice including donor-recipient matching in cadaveric transplantation. Tissue Antigens. 1992 May;39(5):225-35. [ Links ]
17 Garavito EG. Asociación HLA y artritis reumatoidea juvenil: en busca de las bases moleculares dependiente del MHC [tese]. Colombia (CO): Universidad del Norte; 2003.
18 Guo SW, Thompson EA. Performing the exact test of Hardy-Weinberg proportion for multiple alleles. Biometrics.1992 Jun;48(2):361-72. [ Links ]
19 Moraes ME, Fernandez-Viña M, Salatiel I, Tsai S, Moraes JR, Stastny P HLA class II DNA typing in two Brazilian populations. Tissue Antigens. 1993 May;41(5):238-42. [ Links ]
20 Alves-Silva J, Silva SM, Guimarães PEM, Ferreira ACS, Bandelt HJ, Pena SDJ, et al. The ancestry of Brazilian mtDNA lineages. Am J Hum Genet. 2000 Aug;67(2):444-61. [ Links ]
21 Carvalho-Silva DR, Santos FR, Rocha J, Pena SDJ. The phyleogeography of Brazilian Y-chromossomes lineages. Am J Hum Genet. 2001 Jan;68(1):281-6. [ Links ]
22 Polland GA. Influenza Immunization of schoolchildren: can we interrupt community epidemics? Pediatrics. 1999 Jun;103(6 Pt1):1280-2.
23 Parolín ML, Carnese FR. HLA-DRB1 alleles in four Amerindian populations from Argentina and Paraguay. Genet Mol Biol. 2009 Apr;32(2):212-9. [ Links ]
24 Zago MA, Silva Júnior WA, Tavella MH, Santos SE, Guerreiro JF, Figueiredo MS. Interpopulational and intrapopulational genetic diversity of Amerindians as revealed by six variable number of tandem repeats. Hum Hered. 1996 Sep-Oct;46(5):274-89. [ Links ]
25 Desombere I, Willems A, Leroux-Roels G. Response to hepatitis B vaccine: multiple HLA genes are involved. Tissue Antigens. 1998 Jun;51(6):593-604. [ Links ]
26 Nardin EH, Oliveira GA, Calvo-Calle JM, Castro ZR, Nussenzweig RS, Schmeckpeper B, et al. Synthetic malaria peptide vaccine elicits high levels of antibodies in vaccinees of defined HLA genotypes. J Infect Dis. 2000 Nov;182(5):1486-96. Doi:10.1086/315871 [ Links ]
27 Lorenzetti A, Fornazari PA, Bonini-Domingos AC, Souza R, Penhalbel R, Fugikaha E, et al. Mixed Plasmodium falciparum infections and its clinical implications in four areas of the Brazilian Amazon region. Acta Trop. 2008;107(1):8-12. Doi:10.1016/j.actatropica.2008.03.012 [ Links ]
28 Nesse RM, Williams GC. Health and disease. In: Salzano FM, Bortolini MC, editors. The evolution and genetics of Latin American populations. Cambridge: Cambridge University Press; 2002. p. 165-88.
 Correspondence / Correspondência / Correspondencia:
Correspondence / Correspondência / Correspondencia:
Luciane Moreno Storti-Melo
Rua José Duran, no 112.
Bairro: Bosque da Felicidade
CEP: 67030-000
São José do Rio Preto-São Paulo-Brasil
Tel.: +55 (17) 3014-5396
E-mail: stortilu@yahoo.com.br
Received / Recebido em / Recibido en: 2/28/2011
Accepted / Aceito em / Aceito en: 5/17/2011