










Serviços Personalizados
Journal

Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Pan-Amazônica de Saúde
versão impressa ISSN 2176-6215versão On-line ISSN 2176-6223
Rev Pan-Amaz Saude v.2 n.1 Ananindeua mar. 2011
http://dx.doi.org/10.5123/S2176-62232011000100007
ORIGINAL ARTICLE | ARTIGO ORIGINAL | ARTÍCULO ORIGINAL
Identification of a long-standing colony of Proechimys at the Instituto Evandro Chagas, Pará, Brazil, based on cytogenetic information
Identificação de uma colonia de longo prazo de Proechimys no Instituto Evandro Chagas, Pará, Brasil, com base em informações citogenéticas
Identificación de una colonia de largo plazo de Proechimys en el Instituto Evandro Chagas, Pará, Brasil, con base en informaciones citogenéticas
Nelson Antonio Bailão RibeiroI; Julio Cesar PieczarkaII; Manoel do Carmo Pereira SoaresIII; Cleusa Yoshiko NagamachiII
IInstituto Evandro Chagas/SVS/MS, Belém, Pará, Brasil, CAPES Doctor Scholarship on Genetics and Molecular Biology, Brazil
IILaboratório de Citogenética, Instituto de Ciências Biológicas, Universidade Federal do Pará, Belém, Pará, Brasil, Conselho Nacional de Desenvolvimento Científico e Tecnológico Researcher, Brazil
IIIInstituto Evandro Chagas/SVS/MS, Belém, Pará, Brasil
Endereço para correspondência
Correspondence
Dirección para correspondencia
ABSTRACT
The taxonomic classification of the genus Proechimys is complex because many of its species are morphologically similar but chromosomally different, with diploid (2n) values ranging from 14 to 62. The "Seção de Criação e Produção de Animais de Laboratório do Instituto Evandro Chagas" (The Division for Breeding and Production of Laboratory Animals, Instituto Evandro Chagas, Brazil) maintains a Proechimys colony for biomedical research. The colony members have been classified as P. guyannensis, which reportedly has 2n=40 and a fundamental number (FN) = 54. However, using karyotype analysis to aid in their taxonomic classification, we instead observed that a sample of the animals in this colony have 2n = 30 and FN = 56, with a medium-sized submetacentric X chromosome and a small acrocentric Y chromosome. Constitutive heterochromatin was distributed as follows: in the pericentromeric regions of chromosomes 6, 7, 9, 10, 11, 12, 13, 14 and X; on the distal short arms of chromosomes 3, 6, 10 and X; on the distal long arm of chromosome 12; on the long arm of the Y chromosome; and distally on both arms of chromosomes 7, 9 and 11. The nucleolar organizer regions (NORs) are located on the long arm of chromosome 9. This karyotype is consistent with that described previously for P. roberti, but not P. guyannensis, thus demonstrating the importance of using karyotyping for the taxonomic identification of Proechimys.
Keywords: Chromosomes; Proechimys; Rodentia; Karyotyping.
RESUMO
A classificação taxonómica do gênero Proechimys é complexa porque muitas de suas espécies são morfologicamente semelhantes, porém diferentes cromossomicamente, com números de diploide (2n) que variam entre 14 e 62. A Seção de Criação e Produção de Animais de Laboratório do Instituto Evandro Chagas mantém uma colónia de Proechimys para pesquisa biomédica. Os membros da colónia foram classificados como P. guyannensis, que possui 2n = 40 e um número fundamental (NF) = 54. No entanto, ao utilizar a análise do cariótipo para auxiliar em sua classificação taxonómica, observamos que uma amostra dos animais desta colónia possuem 2n = 30 e NF = 56, com um cromossomo X submetacêntrico de tamanho médio e um cromossomo Y acrocêntrico pequeno. A heterocromatina constitutiva foi distribuída da seguinte forma: nas regiões pericentroméricas dos cromossomos 6, 7, 9, 10, 11, 12, 13, 14 e X; na porção distal dos braços curtos dos cromossomos 3, 6, 10 e X; na porção distal do braço longo do cromossomo 12; no braço longo do cromossomo Y; e nas porções distais de ambos os braços dos cromossomos 7, 9 e 11. As regiões organizadoras nucléolos (NORs) localizam-se no braço longo do cromossomo 9. Este cariótipo é consistente com o descrito anteriormente para P. roberti, não para P. guyannensis, o que demonstra a importância do uso de cariotipagem para a identificação taxonómica de Proechimys.
Palavras chave: Cromossomos; Proechimys; Roedores; Cariotipagem.
RESUMEN
La clasificación taxonómica del género Proechimys es compleja porque muchas de sus especies son morfológicamente semejantes, aunque diferentes cromosómicamente, con números de diploide (2n) que varían entre 14 y 62. La Sección de Cría y Producción de Animales de Laboratorio del Instituto Evandro Chagas mantiene una colonia de Proechimys para investigación biomédica. Los miembros de la colonia fueron clasificados como P. guyannensis, que posee 2n = 40 y un número fundamental (NF) = 54. Sin embargo, al utilizar el análisis del cariotipo para auxiliar en su clasificación taxonómica, observamos que una muestra de los animales de esta colonia tenía 2n = 30 y NF = 56, con un cromosoma X submetacéntrico de tamaño mediano y un cromosoma Y acrocéntrico pequeño. La heterocromatina constitutiva fue distribuida de la siguiente forma: en las regiones pericentromérica de los cromosomas 6, 7, 9, 10, 11, 12, 13, 14 y X; en la porción distal de los brazos cortos de los cromosomas 3, 6, 10 y X; en la porción distal del brazo largo del cromosoma 12; en el brazo largo del cromosoma Y; y en las porciones distales de ambos brazos de los cromosomas 7, 9 y 11. Las regiones organizadoras nucleolares (NORs)se localizan en el brazo largo del cromosoma 9. Este cariotipo es consistente con lo descrito anteriormente para P. roberti, no para P. guyannensis, lo que demuestra la importancia del uso de cariotipado para la identificación taxonómica de Proechimys.
Palabras clave: Cromossomos; Proechimys; Roedores; Cariotipificación.
INTRODUCTION
The taxonomic classification of representatives of the family Echimyidae (Hystricognathi - Rodentia) is controversial, with the number of recognized genera ranging from 141 to 162 . Patton and Rogers3 stated that Proechimys is one of the less well-known and more taxonomically complex genera among the Neotropical rodents. There is debate regarding the number of species in this genus because many of these species vary little in their morphological traits. Furthermore, many of the traits that are traditionally used in the systematic classification of mammals, such as fur color, dental enamel pattern, and the number and position of plantar tubercles4,5,6,7, differ in these species based on geography and the age of the animal, further complicating the clear definition of populations and taxa (Figure 1).
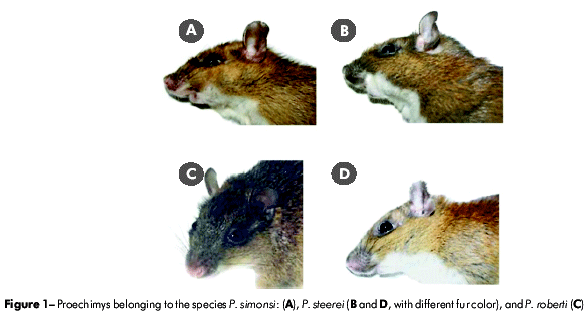
The number of Proechimys species identified in the literature ranges across different authors and papers: Tate8 recognized 46 species, Gardner and Emmons1 identified 32, Ellerman9 reported 21, Moojen5 found 15, and Cabrera10 recognized only 12. Patton11' used bacular and skull data to define 59 species of Proechimys organized into nine groups:
three monotypic groups (decumanus, canicollis, and simonsi) and six polytypic groups (semispinosus, longicaudatus, goeldii, cuvieri, trinitatus and guyannensis). According to this classification, the guyannensis group was composed of the species P. guyannensis, P. cherriei, P. roberti, P. vacilator, P. oris, P. warreni, P. boimensis, P. arescens, P. riparum, and P. arabupu.
These species range from the coastal lands of the Guyanas to the Rio Negro basin and the eastern half of the Amazon basin. Some populations are also found in Goiás and Minas Gerais in Brazil. From this study, the geographic distribution of many species of this genus was defined, such as P. guyannensis, which was found in the left banks of the Amazon River, and P. roberti, which was found in the right banks. This was confirmed by other authors2,12,15,16. However, despite this extensive classification, the number of species in each group remains unclear. More recently, Emmons and Feer12 reported that there are probably no more than 20 or 30 species of Proechimys, whereas Nowak2 found 32 species. Clearly, additional lines of evidence, such as skull structure and karyotype, must also be analyzed concurrently to devise a more precise strategy for identifying the species within Proechimys11.
Based on studies of the sequence of the mitochondrial gene Cytochrome b, Lara et al13 suggested elevating Trinomys, a former subgenus of Proechimys, to the genus level. Leite and Patton14 analyzed the sequences of Cytochrome b, as well as the 12S and 16S genes, which provided support for the proposal of Lara et al13. These data show the potential of molecular studies to elucidate the taxonomy and phylogenetic relationships of this group of rodents.
As Proechimys has huge karyotypic variability, with a diploid (2n) number ranging from 1417 to 6218, cytogenetic analysis appears to be particularly well-suited for resolving the taxonomic problems of this genus. Weksler et al19 used cytogenetic data and other information to evaluate the taxonomic status of the species P. roberti Thomas, 1901 and P. oris Thomas, 1904, and concluded that P. oris is actually a junior synonym of P. roberti. The authors combined their data with that of previous reports to compile a table of the 48 karyotypes described for Proechimys (Table 1), organized according to the species groups proposed by Patton11. The karyotypic characterization of P. roberti was later confirmed by Machado et al20, using samples obtained from Piauí, Tocantins, and Mato Grosso States in Brazil.
MATERIAL AND METHODS
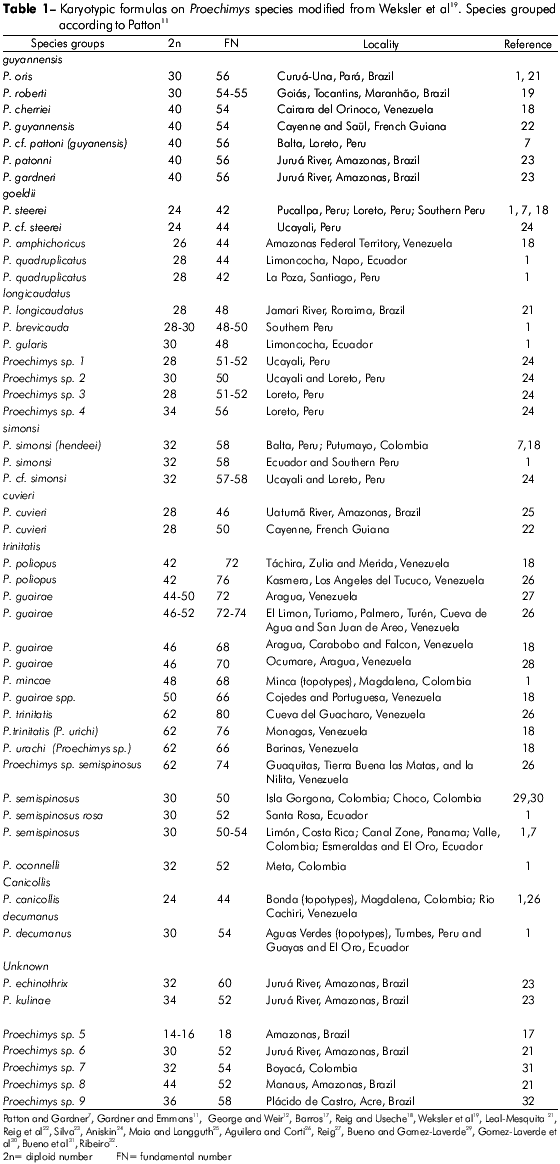
The karyotypes of four animals (two males and two females) of Proechimys belonging to the SACPA-IEC colony were studied. The chromosomes were obtained by direct extraction from bone marrow33 and analyzed by conventional staining using Giemsa, G-banding34, C-banding35 and Ag-NOR staining36. The metaphase chromosomes were photographed under Carl Zeiss III and Axiophot Zeiss microscopes, and karyotypes were mounted based on chromosome morphology, in order of decreasing size. This work was performed in accordance with the Brazilian national ethic rules (Law no. 11.794/08; Arouca Law).
RESULTS
The four animals were found to have 2n = 30 , fundamental number (FN) = 56. The autosomes comprised 13 pairs of biarmed chromosomes, ranging in size from large to small, and one pair of subtelocentrics. The X chromosome was found to be a medium-sized submetacentric chromosome, whereas the Y was a small acrocentric chromosome (Figure 2A). The determination of the chromosome pairs was made by G-banding pattern analysis (Figure 2B).
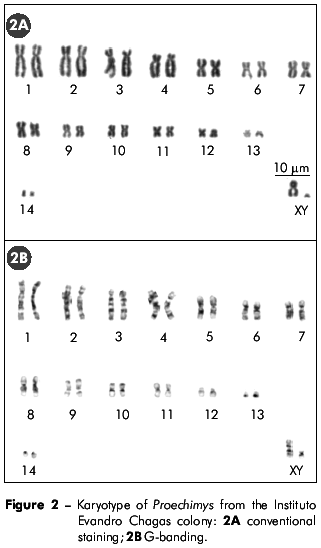
C-banding (Figure 3A) revealed the presence of small blocks of pericentromeric constitutive heterochromatin (CH) in nine autosome pairs (6, 7, 9, 10, 11, 12, 13, 14, and X). CH was also found on the distal short arm of chromosomes 3, 6, 10, and X, on the distal long arm of chromosome 12, on the long arm of the Y chromosome, and distally on both arms of chromosomes 7, 9, and 11.
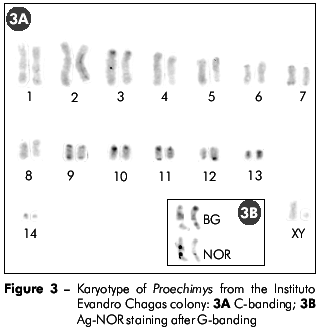
The nucleolar organizer regions (NORs) were located in the interstitial region of the long arm of chromosome 9 (Figure 3B).
DISCUSSION
When the rodent colonies were initially established at the Instituto Evandro Chagas, the Proechimys were classified as P. guyannensis. The published cytogenetic data on this species show karyotypic formulas of 2n=40, FN = 567 for a sample from Balta (Peru), and 2n=40, FN = 5422 for samples from Cayenne and Saul (French Guyana). In sharp contrast, we found that the sample from the SACPA-IEC colony had 2n = 30, FN = 56. Comparison of our data (karyotypic formula, G- and C-banding, and Ag-NOR staining patterns) with those found in the literature for Proechimys shows that our cytogenetic results match those described by Weksler et al19 and Machado et al20 for P. roberti. Additionally, these animals were originally collected in the southwestern region of the Para state. This further suggests that the tested samples must belong to the P. roberti species because previous studies2,12,15,16 demonstrated that P. roberti is found only at the right banks of the Amazon River, where there are no records of P. guyannensis.
Based on these results and those of other studies19,20, the karyotype of the species P. roberti does not have numerical variation, with all tested samples having 2n = 30. However, there are morphological differences involving chromosome 14, the last autosome pair. Populations can be found with a homomorphic, acrocentric pair (FN = 54), a heteromorphic pair with one acrocentric chromosome and a bi-armed chromosome (FN = 54-56), and a homomorphic pair with two bi-armed chromosomes (FN = 56). As previously defined19,20, such chromosomal differences are typical in populations with a defined geographic distribution. This makes them useful to identify populations with biogeographical and phylogenetic similarities.
Weksler et al19 analyzed samples from Goiás and from Pará, Maranhão and Tocantins in the Amazonian region. In the north of Tocantins and Maranhão, they found karyotypes with FN = 54, in which both copies of chromosome 14 were acrocentric. In the other localities, the entire sample had FN = 56, with both copies of chromosome 14 being bi-armed. This is in agreement with the data of Gardner and Emmons1 and Leal-Mesquita21 for samples collected in Curuá-Una, Pará State, and with the data of Barros17 for samples of the Transamazonia Road in Pará. The Proechimys colony from SACPA - IEC originated from the Barros collection17. Thus, our data for this colony are in agreement with Barros. Apart from the homomorphic states of the chromosome 14 pair (acrocentric or bi-armed), Weksler et al19 identified some karyotypes with FN = 56 and FN = 54-56 in the same locality, meaning the chromosome 14 pair is heteromorphic. According to the authors, this is a consequence of a pericentric inversion in one of the homologues. These karyotypic patterns (NF = 56 and NF =54-56) were found in populations of the Cavalcante Farm, northeastern Goiás, and Primavera Farm, northeastern Pará. Machado et al20 also analyzed karyotypes of P. roberti from six regions of three Brazilian states where they found karyotypes with FN = 56 and bi-armed chromosome 14 homologues: the Ecological Station at Uruçuí-Una, Piauí; Parana and Peixe, Tocantins; Cláudia, Gaúcha do Norte and Vila Rica, Mato Grosso.
From the geographic and karyotypic analyses of P. roberti, the existence of two populations can be noted: a western population, in Pará, north of Tocantins, Goiás and Mato Grosso, with FN = 56 and a homomorphic, bi-armed chromosome 14 homolog pair; and an eastern population, located in Maranhão, south of Tocantins, with FN = 54 and an acrocentric chromosome 14 homolog pair. However, Weksler et al19 found karyotypes with FN = 54-56 (with heteromorphic copies of chromosome 14) in populations from the Cavalcante farm, northeastern Goiás, and Primavera Farm, northeastern Pará. However, it is possible that these are hybrid populations, resulting from the mix of the homomorphic groups (one-armed in the east and bi-armed in the west), as the heteromorphic populations are observed in the contact region of both the homomorphic types (northeast of Para and Goiás). This region may thus constitute a potential hybridization zone.
CONCLUSION
This mistake in classification is very understandable given all the problems related to classifying members of genus Proechimys, such as the relative lack of morphological variability among species and the intrapopulational variations in some traits typically used for mammal systematics. At the time of the colony's initial classification four decades ago, taxonomic identification was made exclusively using morphology.
Since many biomedical studies deal with the species-specific relationships among vertebrate reservoirs and pathogenic species (viruses, bacteria, fungi, protozoa, helminthes, etc.) that have been established over thousands of years of co-evolution, with different vertebrate species having different reactions to pathogens, it is very important that model animals be correctly characterized. Due to the morphological similarities among species of Proechimys, these rodents should be accurately identified using techniques beyond morphological examination. Here, we show that cytogenetic studies, in conjunction with other data, are very useful tools for the precise definition of members of Proechimys.
By comparing the cytogenetic data of the sample of P. roberti from SACPA-IEC with published samples, we concluded that the SACPA-IEC sample belongs to the largest populational group of this species, with FN = 56 and a geographical distribution ranging from Pará to Mato Grosso. This suggests that the sample shares a similar evolutionary origin with that group and therefore must have the same genetic and biological properties that allow its distribution and survival in a forest environment characterized by a particular complement of pathogenic species. The SACPA-IEC P. roberti samples may thus represent an ideal model system in which to study species-specific relationships with pathogens.
ACKNOWLEDGMENTS
We thank Dr. Regina Barros (UFPa) for her expertise in the field of cytogenetics, Doctor Reinaldo Carvalho (chairman of the SACPA-IEC at the time of this study) for permission to collect samples, the Instituto Evandro Chagas and Universidade Federal do Pará for the use of laboratory facilities, and CNPq and CAPES for financial support.
FINANCIAL SUPPORT
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Conselho Nacional de Desenvolvimento Científico e Tecnológico, Universidade Federal do Pará, Instituto Evandro Chagas.
REFERENCES
1 Gardner AL, Emmons LH. Species groups in Proechimys (Rodentia, Echimyidae) as indicated by kariology and bullar morfology. J mammal. 1984;65:10-25.
2 Nowak RM. Walker's Mammals of the World. 5th. ed. Baltimore: The Johns Hopkins University Press; 1999. 642 p.
3 Patton JL, Rogers A. Systematic implications of non-geographic variation in the spiny rats genus Proechimys (Echimyidae). Z Sdugetierkd. 1983;48:363-70.
4 Thomas O. The Godman-Thomas expedition to Peru. VII. The mammals of the Rio Ucayali. Ann Mag Nat Hist Ser. 1928;6:294-302.
5 Moojen J. Speciation in the Brazilian spiny rats (genus Proechimys, family Echimyidae). Univ Kans Pubis Mus Nat Hist. 1948;1:301-406.
6 Hershkovitz P. Mammals of norther Colombia. Preliminary report no 2: Spiny Rats (Echimyidae), with suplemental notes on related forms. Proceedings of the United States National Museum. 1948;97:125-40. [Link]
7 Patton JL, Gardner LA. Notes on the systematics of Proechimys (Rodentia: Echimyidae), with emphasis on Peruvian forms. Occasional papers of the Museum of Zoology. 1972;44:1-30.
8 Tate GHH. The taxonomy of the genera of Neotropical hystricoid rodents. Bull Amer Mus Nat Hist. 1935;68:295-447.
9 Ellerman JR. The families and genera of living rodents. London, British: Museum Natural History; 1940. 689 p.
10 Cabrera A. Catalogo de los mamíferos de América del Sur. Mus Argent Ci Nat "Bernardino Rivadavia". 1961;4(2):309-732.
11 Patton JL. Species groups of spiny rats, genus Proechimys (Rodenta: Echimyidae). Fieldiana Zool. 1987;39:305-45.
12 Emmons LH, Feer F. Neotropical rainforest mammals: a field guide. 2nd. ed. Chicago: University of Chicago; 1997.307 p.
13 Lara MC, Patton JL, Silva MNF. The simultaneous Diversification of South American Echimyid Rodents (Hystricognathi) Basead on Complete Cytochrome b Sequences. Mol Phylogenet Evol. 1996 Apr;5(2):403-13. Doi:![]() 10.1006/mpev.1996.0035 [Link]
10.1006/mpev.1996.0035 [Link]
14 Leite YLR, Patton JL. Evolution of South American spiny rats (Rodentia, Echimyidae): the star-phylogeny hypothesis revisited. Mol Phylogenet Evol. 2002 Dec;25(3):455-64. Doi:10.1016/S1055-7903(02)00279-8 [Link]
15 Wilson DE, Reeder DM. Mammal species of the world. A taxonomic and geographic reference. 2nd. ed. Washington: Smithsonian Inst. Press; 1993. 1114 p.
16 Eisenberg JF, Redford KH. Mammals of the Neotropics. Central Neotropics. Chicago: University of Chicago Press; 1999. s.n.
17 Barros RMS. Variabilidade cromossômica em Proechimys e Oryzomys (Rodentia) da Amazônia [Tese]. São Paulo (SP): Universidade de São Paulo, Instituto de Biociências; 1978.
18 Reig AO, Useche M. Diversidad cariotípica y sistemática en poblaciones venezolanas de Proechimys (Rodentia: Echimyidae), com datos adicionales sobre poblaciones de Peru y Colombia. Acta Cient Venz. 1976;27(3):132-40.
19 Weksler M, Bonvicino CR, Otazu I, Silva Junior JSS. Status of Proechimys roberti and P-oris (Rodentia: Echimyidae) from eastern Amazonia and central Brazil. J Mammal. 2001;82(1):109-22. Doi:10.1644/1545-1542(2001)082<0109:SOPRAP>2.0.CO;2 [Link]
20 Machado T, Silva MJJ, Leal-Mesquita ER, Carmignotto AP, Yonenaga-Yassuda Y. Nine karyomorphs for spiny rats of the genus Proechimys (Echimyidae, Rodentia) from North and Central Brazil. Genet mol Biol. 2005;28:682-92. Doi:10.1590/S1415-47572005000500007 [Link]
21 LLeal-Mesquita ERRBP. Estudos Citogenéticos em dez espécies de roedores brasileiros da família Echimyidae [Tese]. São Paulo (SP): Universidade de São Paulo; 1991.
22 Reig OA, Trainer M, Barros MA. Sur l'i'dentification chromosomique de Proechimys guyannensis (E. Geoffroy, 1803) et de Proechimys cuvieri Petter, 1978 (Rodentia : Echimyidae). Mammalia. 1979;43:501-5.
23 Silva MNF. Four new species of spiny rats of the genus Proechimys (Rodentia: Echimyidae) from the western Amazon of Brasil. Proc Biol Soc Wash. 1998;111(2):436-71.
24 Aniskin VM. Evolutionary cytogenetics of spiny rats from Peruvian Amazonia (Rodentia, Echimyidae). In: Mlekopitayushchie Peruanskoi Amazonii. Moskow, Russia: V. E. Sokolov; 1994. 62-82 p.
25 Maia V, Langguth A. Constitutive heterochromatin polymorphism and NORs in Proechimys cuvieri Petter, (Rodentia, Echimyidae). Rev bras genet. 1993;16(1):145-54. [Link]
26 Aguilera M, Corti M. Craniometric differentiation and chromosomal speciation of the genus Proechimys (Rodentia: Echimyidae). Z Sãugetierkunde. 1994;59:366-77.
27 Reig OA. Karyotypic repatterning as one triggering factor in explosive speciation. In: Evolutionary biology of transient unstable populations. Berlin, Germany: Springer-Verlag; 1989. 246-89 p.
28 George W, Weir BJ. A note on the karyotype of Proechimys guairae (Rodentia: Hystricomorpha). Mammalia. 1973;37:330-2.
29 Bueno ML, Gomez- Laverde M. Variacion Heterocromática em Proechimys semispinosus (Rodentia: Echimyidae) de la Region Pacifica Colombiana. Caldasia. 1993;7(2):333-40.
30 Gomez-Laverde M, Bueno ML, Cadena A. Poblaciones de ratas (Proechimys semispinosus) (Rodentia: Echimyidae). In: Aguirre J, Rangel O, editors. Biota y Ecosistema de Gorgona y Gorgonilla. Bogotá, Colombia: Editorial Presencia; 1990. 244-51 p.
31 Bueno ML, Gomez-Laverde M, Morales A. Caracterización cariológica de Proechimys sp (Rodentia: Echimyidae) de una colônia experimental. Biomédica. 1989;9:13-22.
32 Ribeiro NAB. Análises Cromossômicas e Filogenia de Roedores do Gênero Proechimys (Echimyidae, Rodentia) [Tese]. Belém (PA): Universidade Federal do Pará, Centro de Ciências Biológicas; 2006. [Link]
33 Ford CE, Hamerton JL. A colchicine, hypotonic -citrate, squash sequence for mammalian chromosomes. Stain Technol. 1956;31(6):247-51.
34 Seabright M. "The use of proteolitic enzymes for the mapping of structural rearrangements of man". Chromosoma. 1971;36:204-10.
35 Sumner AT. A simple technique for demosntrating cen tromeric heterocromatin. Exp cell Res. 1972;75:304-6.
36 Howell WM, Black DA. Controlled silver- staining of nucleolar organizer regions whith protective colloidal developer: a 1-step method. Experientia. 1980;36(8):1014-5.
 Correspondência /Correspondence / Correspondencia:
Correspondência /Correspondence / Correspondencia:
Nelson Antonio Bailão Ribeiro
Rua dos Pariquís
Alameda Gonçalo Duarte no 56
Bairro: Jurunas
CEP:66030-040
Belém-Pará-Brasil
E-mail:nelsonribeiro@iec.pa.gov.br
Recebido em: 9/9/2010
Aprovado em: 4/5/2011
