










Serviços Personalizados
Journal

Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Pan-Amazônica de Saúde
versão impressa ISSN 2176-6215versão On-line ISSN 2176-6223
Rev Pan-Amaz Saude v.2 n.2 Ananindeua jun. 2011
http://dx.doi.org/10.5123/S2176-62232011000200009
COMMUNICATION | COMUNICAÇÃO | COMUNICACIÓN
Susceptibility of a continuous murine cell line (GRX) to viral infection
Suscetibilidade de uma linhagem celular murina contínua (GRX) à infecção viral
Susceptibilidad de un linaje celular murino continuo (GRX) a la infección viral
Maria Liz GamarraI; Maria Carolina Maciel AlbuquerqueI; Anderson Junger TeodoroII; Renata Brum MartucciIII; Radovan BorojevicII; Fernando Portela CâmaraI; Maria Teresa Villela RomanosI; Norma SantosI
IDepartamento de Virologia, Instituto de Microbiologia Paulo de Góes, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Rio de Janeiro, Brasil
IIDepartamento de Histologia, Instituto de Ciências Biológicas, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Rio de Janeiro, Brasil
IIIDepartamento de Nutrição Aplicada, Instituto de Nutrição, Universidade Estadual do Rio de Janeiro, Rio de Janeiro, Brasil
Endereço para correspondência
Correspondence
Dirección para correspondencia
ABSTRACT
The ability of a murine cell line (GRX) to support viral replication was evaluated. GRX cell cultures were infected with different DNA or RNA viruses. It was observed that the GRX cell line is susceptible to the replication of Herpes simplex virus types 1 and 2 (HSV-1 and HSV-2), Mayaro virus (MAY), Sindbis virus (SIN), and West equine encephalitis virus (WEE), and can be used as substrate for viral replication studies. Viral replication induced cytopathic effect (CPE) 24-48 h post-infection. The GRX cells yielded infectious virus titers between 102.4 TCID50 (Tissue Culture Infectious Dose50) /25 µL and 105.4 TCID50/25 µL in the first viral passage. These results demonstrate that GRX cells efficiently sustain viralreplication and therefore can be used as a valuable tool in the virology laboratory.
Keywords: Cell culture technique; Hepatic Stellate Cells (GRX); Virus replication.
RESUMO
Este estudo avaliou a capacidade de uma linhagem celular murina (GRX) de realizar a replicação viral. Culturas de células GRX foram infectadas com diferentes vírus DNA e RNA. Foi observado que a linhagem celular GRX é suscetível à replicação dos vírus Herpes simplex tipos 1 e 2 (HSV-1 e HSV-2), Mayaro (MAY), Sindbis (SIN) e vírus da encefalite equina do oeste (WEE) e pode ser utilizada como suporte para estudos sobre replicação viral. A replicação viral induziu o efeito citopático 24 a 48 h pós-infecção. As células GRX produziram titulações de vírus infecciosos entre 102.4 TCID50 (dose infecciosa de cultura de tecido50)/25 µL e 105.4 TCID50/25 µL na primeira passagem viral. Esses resultados demonstram que as células GRX sustentam, de forma eficiente, a replicação viral e, portanto, podem ser utilizadas como uma ferramenta valiosa para estudos laboratoriais sobre virologia.
Palavras chave: Técnica de cultura de células; Células Estreladas do Fígado (GRX); Replicação Viral.
RESUMEN
Este estudio evaluó la capacidad de un linaje celular murino (GRX) de realizar la replicación viral. Culturas de células GRX fueron infectadas con diferentes virus ADN y ARN. Se observó que el linaje celular GRX es susceptible a la replicación de los virus Herpes simplex tipos 1 y 2 (HSV-1 y HSV-2), Mayaro (MAY), Sindbis (SIN) y al virus de la encefalitis equina del oeste (WEE) y puede utilizarse como soporte para estudios sobre replicación viral. La replicación viral indujo el efecto citopático 24 a 48 h pos-infección. Las células GRX produjeron titulaciones de virus infecciosos entre 102.4 TCID50 (dosis infecciosa de cultura de te¡¡do50)/25 µL y 105.4 TCID50/25 µL en el primer pasaje viral. Estos resultados demuestran que las células GRX sostienen, de forma eficiente, la replicación viral y, por lo tanto, pueden ser utilizadas como una valiosa herramienta para estudios de laboratorio sobre virología.
Palabras clave: Técnicas de Cultivo de Célula; Células Estrelladas Hepáticas (GRX); Replicación Viral.
During the last two decades, the application of molecular and serological methodologies has had a major impact on the detection of novel viruses, yet viral isolation remains the gold standard method for the identification and characterization of their biological and biochemical properties. Cell culture is still the most common method for propagation of viruses. Cell culture methods are also used for vaccine production and biochemical and molecular biological studies of viral replication1. In 1985, GRX, a continuous cell line representative of hepatic stellate cells, was established from fibro-granulomatous lesions induced in mouse liver by schistosomal infection2, and its biological and biochemical characteristics were determined3,4,5. It is a highly proliferative cell line showing fibroblastic morphology with a stellate, polygonal or elongated shape. When the monolayers are confluent, the cells gather in a distinct "hills and valleys" pattern of growth2. The ability of the GRX cell line to support viral replication has never been shown; therefore, the primary goal of this work was to demonstrate the usefulness of the GRX cell line for propagation of selected viruses.
The GRX cells were obtained from the Banco de Células do Estado do Rio de Janeiro, Rio de Janeiro, Brazil. The African green monkey kidney (Vero), rhesus monkey kidney (MA-104), and human epithelial (HEp-2) cell lines were obtained from stocks used for routine viral isolation in the Virology Department of the Microbiology Institute, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil. All cell lines were cultured in Minimum Essential Medium (MEM) supplemented with 5% (HEp-2 cells) or 10% (Vero, MA-104, and GRX cells) heat-inactivated fetal bovine serum (FBS), 2 mM L-glutamine, 50 µg/mL gentamicin, 2.5 µg/mL fungizone, sodium bicarbonate at 0.25%, and 10 mM HEPES. Cell cultures were incubated at 37o C with 5% CO2.
The virus strains used in this paper were obtained from the reference collection of the Virology Department of the Microbiology Institute, Universidade Federal do Rio de Janeiro. Herpes simplex virus type 1 (HSV-1) and type 2 (HSV-2), Sindbis virus (SIN), Mayaro virus (MAY), and West equine encephalitis virus (WEE) were maintained in Vero cells; Adenovirus (AdV) serotypes 2, 19, 40, and 41 were maintained in HEp-2 cells; and Rotavirus (RV) strain SA-11 was maintained in MA-104 cells.
To propagate the viruses, cell monolayers were prepared in 48-well plates; each viral suspension was inoculated in triplicate. The inoculated plates were incubated for 1 h at 37o C. Subsequently, the inocula were replaced with fresh MEM without FBS (maintenance medium). For rotavirus, trypsin was added to the inoculum at a final concentration of 10 µg/mL, followed by incubation at 37o C for 30 min. Then the inoculum was added to the cell monolayer with maintenance medium at a final trypsin concentration of 5 µg/mL. All infected cell cultures were incubated at 37o C with 5% CO2. The cells were monitored daily for the development of viral cytopathic effect (CPE) using an inverted microscope. When approximately 75% of the cells in the culture showed CPE or after seven days of incubation in the absence of CPE, the infected cell cultures were subjected to three freeze-thaw cycles. Afterwards, the mixtures of cell lysates and supernatants were collected and stored at -70o C for future use. Each virus strain was subjected to three serial passages in GRX cells. The mixtures of supernatant and cell lysate (viral suspension) from each of the three passages were subjected to titration by the endpoint dilution method to quantify viral infectivity. Logarithmic dilutions of 25 µL of the viral suspension were inoculated onto confluent monolayers in 96-well plates (six wells/dilution). Titers were calculated according to the Reed and Muench method and expressed in log10 TCID50 (Tissue Culture Infectious Dose50)/25 µL6.
In 24-well plates, GRX cells at a density of 4.2 x 105 cells/mL and Vero cells at a density of 1.2 x 106 cells/mL were plated in MEM supplemented with 10% FBS and infected with 100 µL of viral suspension (MAY, SIN or WEE) at a multiplicity of infection (MOI) of 1.0, 0.5, and 0.1. The infected cell cultures were incubated at 37o C with 5% CO2 for 48 h. After that period, the viral suspension was collected and titrated as described above. All experiments were performed in triplicate.
Statistical analysis was performed using analysis of variance (ANOVA) to determine the least significant difference at p = 0.05. The analysis was performed with Minitab® for Windows, Release 14.0 (Minitab Inc., State College, PA, USA). The term "significance" (statistically significant) in the text means p≤0.05.
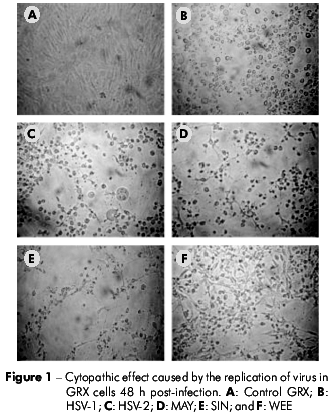
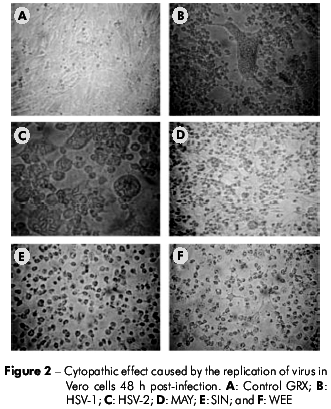
GRX cells were susceptible to infection with HSV-1, HSV-2, SIN, and WEE, producing visible CPE in 48 h. MAY induced CPE in 24 h. In contrast, GRX cells did not support detectable viral replication when inoculated with AdV or RV (Table 1). The CPE observed for MAY, SIN, and WEE on GRX cells was cell pyknosis followed by cytoplasm fragmentation and detachment from the monolayer, which was similar to the CPE observed in Vero cells. For HSV-1 and HSV-2, the CPE observed was rounding and refringence of the cells (Figures 1 and 2).
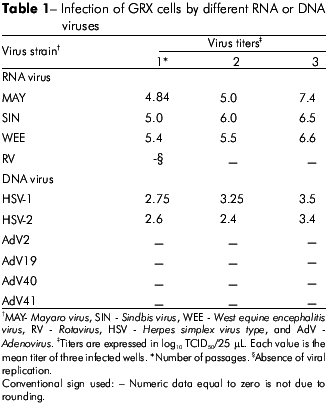
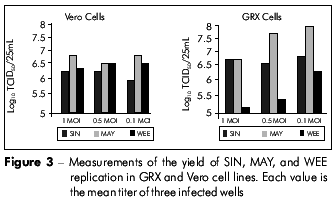
To demonstrate the efficiency of viral replication, we measured the viral yield in GRX cells compared to that of Vero cells after inoculation with virus at different MOIs. We observed no statistically significant difference between the two cell lines, indicating that viral replication occurs efficiently in both systems (Figure 3). For Vero cells, no significant difference was observed when the viruses were inoculated at different MOIs. For GRX cells, although no significant differences in viral yield were observed for SIN and MAY at different MOIs, we observed some variance in the yield of MAY replication when these viruses were inoculated at a low MOI. Conversely, there was a significant difference (p=0.016) in the viral yield for WEE when different MOIs were used (Figure 3).
Although several cell lines have been used for viral isolation, there are still a significant number of viruses that cannot be propagated in the laboratory. Therefore, the development of a new cell line susceptible to viral replication engenders new alternatives for the cultivation of such viruses. The search for new alternatives has been intensified lately, mainly due to the emergence of novel viruses, such as the human metapneumovirus7 and the new human coronaviruses (SARS-CoV, HCoV-NL63, HCoV-HKU1)8,9,10, or to the necessity of viral isolation for biochemical or molecular studies as in the case of norovirus11, hepatitis B virus12, and hepatitis C virus13. Consequently, studies have been published evaluating the utility of several cell lines for virus isolation14,15,16.
The GRX cell line was first described in 19852; however, the usefulness of these cells to viral replication is yet to be determined. In this study, we evaluated the ability of the GRX cell line to support the propagation of several viruses and its usefulness as a tool for virology laboratories. Generally, the determination of the susceptibility of a cell line to viral replication can be achieved by observing the characteristic CPE followed by viral titration or by using alternative methodologies, such as a hemagglutination test, immunofluorescence or the detection of the viral genome by molecular methods14,15,16. In this study, we used CPE observations and viral titration as the parameters for demonstrating cell susceptibility. All the viruses used in this study produced visible CPE in other cell lines; hence, it would not be advantageous to establish a new cell line for those viruses that require alternative methodologies to demonstrate virus propagation in culture.
The alphaviruses WEE, MAY, and SIN infect a large range of hosts in nature, replicating in mammals, birds, arthropods, and amphibians, and these viruses can be propagated in vitro in a great variety of cell lines17,18,19. Inoculation of the GRX cells with these viruses produced CPE in 24-48 h. High viral titers were obtained starting with the first passage. The titers increased progressively with consecutive passages, demonstrating the production of infectious virus particles. The GRX cells provided a substrate for viral propagation as efficiently as the Vero cells, which are regularly used for the propagation of alphaviruses.
Different cell lines can be used for the isolation of HSVs20. HSV-1 and HSV-2 were successfully propagated in GRX cells at 48 h post-inoculation, showing that although the viral titers obtained were lower than those from Vero cells, GRX cells can be efficiently used as a culture system for these viruses.
Interestingly, in the present study all the viruses that successfully propagated in the GRX cells are enveloped (Alphaviruses) as opposed to non-enveloped viruses, such as adenovirus and rotavirus. One possible explanation for this phenomenon could be the specificity of the viral receptor. Another explanation could be the presence of cellular enzymes required for viral replication. More studies need to be carried out to clarify this issue.
The results showed that the GRX cell line exhibited high susceptibility to different viruses, yielding high titers. Virus-induced CPE was observed between 24 and 48 h, depending on the virus used. The measurement of viral yield demonstrated that the titers progressively increased with the passages, indicating that the GRX cells can efficiently sustain viral replication. The GRX cell line is highly susceptible to HSV-1, HSV-2, MAY, SIN, and WEE, and it can be used as a tool for viral isolation and biochemical studies.
Although there are numerous cell lines widely used in virology to propagate viruses, the establishment of a new cell system always opens new possibilities for exploring the biology and biochemistry of viral infections. The GRX cell line was established over in the earliest 1980s, a nd its biological and biochemical characteristics have been determined. However, its ability to support viral replication has not been clearly demonstrated. The primary goal of this work was to demonstrate the utility of the GRX cell line for viral propagation. For comparison, we worked initially with viruses that already have an efficient cell system. After establishing the GRX cell line's susceptibility to particular viruses, we can continue to study its ability to support the growth of viruses that have not yet been propagated in vitro, such as hepatitis C virus and human bocavirus.
In conclusion, (i) we were able to fulfill our primary goal of demonstrating the susceptibility of the GRX cell line to virus infection, (ii) we have shown that this cell line is a valuable virological tool, and (iii) our results open the possibility of using the GRX cell line for viruses not previously propagated in vitro by us or by any other laboratory interested in such studies.
ACKNOWLEDGEMENTS
We thank Soluza dos Santos Gonçalves for technical assistance.
FINANCIAL SUPPORT
This study was supported in part by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), and Fundação Carlos Chagas de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), Brazil.
REFERENCES
1 Olivo PD. Transgenic cell lines for detection of animal viruses. Clin Microbiol Rev. 1996 Jul;9(3):321-34. [Link]
2 Borojevic R, Monteiro ANA, Vinhas SA, Domont GB, Mourão PAS, Emonard H, et al. Establishment of a continuous cell line from fibrotic schistosomal granulomas in mice livers. In Vitro Cell Dev Biol. 1985 Jul;21(7):382-90. [Link]
3 Fortuna VA, Trugo LC, Borojevic R. Acyl-CoA: retinol acyltransferase (ARAT) and lecithin: reti no l acyltransferase (LRAT) activation during the lipocyte phenotype induction in hepatic stellate cells. J Nutr Biochem. 2001 Nov;12(11):610-21. Doi:10.1016/S0955-2863(01)00179-6 [Link]
4 Guma FCR, Mello TG, Mermelstein CS, Fortuna V, Wofchuk ST, Gottfried C, et al. Intermediate filaments modulation in an in vitro model of the hepatic stellate cell activation or conversion into the lipocyte phenotype. Biochem Cell Biol. 2001;79(4):409-17. [Link]
5 Martucci RB, Ziulkoski AL, Fortuna VA, Guaragna RM, Guma FCR, Trugo LC, et al. β-carotene storage, convertion to retinoic acid, and induction of the lipocyte in hepatic stellate cells. J Cell Biochem. 2004 May;92(2):414-23. DOI:10.1002/jcb.20073 [Link]
6 Reed LJ, Muench H. A simple method of estimating fifty per cent endpoints. Am J Hyg. 1938 May;27(3):493-7.
7 Van den Hoogen BG, Jong JC, Groen J, Huiken T, Groot R, Fouchier RAM, et al. A newly discovered human pneumovirus isolated from young children with respiratory tract disease. Nat Med. 2001 Jun;7(6):719-24. Doi:10.1038/89098 [Link]
8 Ksiazek T, Erdman D, Goldsmith CS, Zaki SR, Peret T, Emery S, et al. A novel coronavirus associated with severe acute respiratory syndrome. N Engl J Med. 2003 May;348(20):1953-66. [Link]
9 Van der Hoek L, Pyrc K, Jebbink MF, Vermeulen-Oost W, Berkhout RJM, Wolthers KC, et al. Identification of a new coronavirus. Nat Med. 2004 Apr;10(4):368-73. [Link]
10 Woo PCY, Lau SKP, Chu CM, Chan KH, Tsoi HW, Huang Y, et al. Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumonia. J Virol. 2005 Jan;79(2):884-95. Doi:10.1128/JVI.79.2.884-895.2005 [Link]
11 Duizer E, Schwab KJ, Neil FH, Atmar RL, Koopmans MP, Estes MK. Laboratory effort to cultivate norovirus. J Gen Virol. 2004 Jan;85(Pt 1):79-87. [Link]
12 Guha C, Mohan S, Roy-Chowdhury N, Roy-Chowdhury J. Cell culture and animal models of viral hepatitis. Part I: hepatitis B. Lab Anim (NY). 2004 Jul-Aug;33(7):37-46. [Link]
13 Durantel D, Zoulim F. Going towards more relevant cell culture models to study the in vitro replication of serum-derived hepatitis C virus and virus/host cell interactions? J Hepatol. 2007 Jan;46(1):1-5.
14 El-Awady MK, Tabll AA, El-Abd YS, Bahgat MM, Shoeb HA, Youssef SS, et al. HepG2 cells support viral replication and gene expression of hepatitis C virus genotype 4 in vitro. World J Gastroenterol. 2006 Aug;12(30):4836-42. [Link]
15 Ingram RE, Fenwick F, McGuckin R, Tefari A, Taylor C, Toms GL. Detection of human metapneumovirus in respiratory secretions by reverse-transcriptase polymerase chain reaction, indirect immunofluorescence, and virus isolation in human bronchial epithelial cells. J Med Virol. 2006 Sep;78(9):1223-31. DOI:10.1002/jmv.20685 [Link]
16 Qin QW, Wu TH, Jia TL, Hegde A, Zhang RQ. Development and characterization of a new tropical marine fish cell line from grouper, Epinephelus coioides susceptible to iridovirus and nodavirus. J Virol Meth. 2006 Jan;131(1):58-64. Doi:10.1016/J.Jviromet.2005.07.009 [Link]
17 Bello FJ, Rodríguez JA, Escovar J, Olano VA, Morales A, González M, et al. A new continuous cell line from the mosquito Psorophora confinnis (Diptera: Culicidae) and its susceptibility to infections with some arboviruses. Mem Inst Oswaldo Cruz. 2001 Aug;96(6):865-73. http://dx.doi.org/10.1590/S0074-02762001000600022 [Link]
18 Ferreira DF, Santo MPE, Rebello MA, Rebello MCS. Weak bases affect late stages of Mayaro virus replication cycle in vertebrate cell. J Med Microbiol. 2000 Apr;49(4):313-8. [Link]
19 Weingartl HM, Sabara M, Pasick J, Van Moorlehem E, Babiuk L. Continuous porcine cell line developed from alveolar macrophages: partial characterization and virus susceptibility. J Virol Meth. 2002 Jul;104(2):203-16. Doi:10.1016/S0166-0934(02)00085-X [Link]
20 Ustaçelebi S. Diagnosis of herpes simplex virus infections. J Clin Virol. 2001 Jun;21(3):255-9.
 Correspondence / Correspondência / Correspondencia:
Correspondence / Correspondência / Correspondencia:
Norma Santos
Departamento de Virologia,
Instituto de Microbiologia Paulo de Góes
Universidade Federal do Rio de Janeiro
Cidade Universitária, CCS - Bl. I, Ilha do Fundão
CEP: 21.941-590
Rio de Janeiro-Rio de Janeiro-Brasil
Tel: + 55 (21) 2560-8344 Fax + 55 (21) 2560-88028
E-mail:nsantos@micro.ufrj.br
Received / Recebido em / Recibido en: 3/5/2011
Accepted / Aceito em / Aceito en: 16/9/2011
