










Serviços Personalizados
Journal

Artigo
 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Pan-Amazônica de Saúde
versão impressa ISSN 2176-6215versão On-line ISSN 2176-6223
Rev Pan-Amaz Saude v.4 n.3 Ananindeua set. 2013
http://dx.doi.org/10.5123/S2176-62232013000300004
ARTIGO ORIGINAL | ORIGINAL ARTICLE | ARTÍCULO ORIGINAL
Busca por Hepacivirus semelhante ao vírus da hepatite C, em primatas não humanos
Searching for Hepacivirus related to hepatitis C virus in nonhuman primates
Busca por Hepacivirus parecido al virus de la hepatitis C, en primates no humanos
Gustavo Moraes Holanda; Manoel do Carmo Pereira Soares; Nelson Antonio Bailão Ribeiro
Seção de Hepatologia, Instituto Evandro Chagas/SVS/MS, Belém, Pará, Brasil
Endereço para correspondência
Correspondence
Dirección para correspondencia
RESUMO
O gênero viral Hepacivirus é constituído pelo vírus da hepatite C (VHC), porém há proposta de inclusão de outros vírus recentemente encontrados em cães, cavalos e roedores e do vírus GB B. O VHC, ou algum outro vírus filogeneticamente relacionado a ele, ainda não foi identificado em primatas não humanos silvestres, diferentemente de vírus semelhantes ao vírus da hepatite B. As infecções causadas pelo VHC em humanos são graves existindo, assim, a necessidade de uma busca, em outros animais, por Hepacivirus, visto que eles poderão cruzar a barreira evolutiva, causando infecção em seres humanos. O objetivo deste trabalho foi tentar localizar a presença de Hepacivirus em primatas não humanos. Foram desenhados iniciadores, os quais se ligam na região IRES do VHC, obtendo um produto final de 188 pb. Foram analisadas 1.054 amostras, pertencentes a 36 espécies de primatas não humanos das quais apenas 12 apresentaram bandas definidas e, após sequenciamento, apresentaram negatividade para Hepacivirus. Em análises evolutivas observou-se que o vírus GB C ocupa a mais distante posição filogenética em relação ao VHC, enquanto a mais próxima é ocupada pelo vírus GB B. Apesar da não identificação de Hepacivirus semelhantes ao VHC em primatas não humanos, não se pode afirmar que o mesmo não exista. Estudos envolvendo outras espécies, como os vírus GBs e os possíveis novos Hepacivirus podem solucionar o processo evolutivo que possibilitou o VHC infectar seres humanos, pois estes parecem ser o elo entre o VHC e espécies virais encontradas em animais silvestres.
Palavras-chave: Hepacivirus; Pegivirus; Vírus GB A; Vírus GB B; Vírus GB C.
ABSTRACT
Hepacivirus genus comprises hepatitis C virus (HCV), but a proposal to include other viruses, originated from dogs, horses, rodents, and GB virus B (GBV-B), is being made. HCV, or any other virus phylogenetically related to it, has not been identified in wild nonhuman primates, differently from HBV-like viruses. HCV infections in humans are severe, therefore a search for Hepacivirus in other animals is needed for they can cross the evolutionary barrier causing infections in humans. This study tried to identify a nonhuman primate origin for Hepacivirus. Primers have been designed, which bind on HCV IRES region, resulting a final product of 188 bp. A total of 1,054 samples collected from 36 nonhuman primate species were analyzed, of which just 12 presented defined bands and were negative for Hepacivirus after sequencing. In evolutionary analysis, GB virus C in a phylogenetic position is the most distant from HCV, while GBV-B is the nearest. Although HCV-related Hepacivirus is not found in nonhuman primates, its existence cannot be denied. Studies involving other species, such as GB viruses and possible new Hepacivirus, can solve the evolutionary process that has allowed HCV infect humans, since they seem to be the link between HCV and viral species found in wild animals.
Keywords: Hepacivirus; Pegivirus; GB virus A; GB virus B; GB virus C.
RESUMEN
El género viral Hepacivirus está constituido por el virus de la hepatitis C (VHC), pero existe una propuesta de inclusión de otros virus recientemente encontrados en perros, caballos y roedores y por el virus GB B. El VHC, o algún otro virus filogenéticamente relacionado con él, todavía no han sido identificados en primates no humanos silvestres, diferentemente a virus semejantes al virus de la hepatitis B. Las infecciones causadas por el VHC en humanos son graves y por eso existe la necesidad de una búsqueda, en otros animales, de Hepacivirus, visto que estos podrían cruzar la barrera evolutiva, causando infección en seres humanos. El objetivo de este trabajo fue el de intentar localizar la presencia de Hepacivirus en primates no humanos. Se diseñaron iniciadores que se conectan en la región IRES del VHC, obteniendo un producto final de 188 pb. Fueron analizadas 1.054 muestras, pertenecientes a 36 especies de primates no humanos de la cuales apenas 12 presentaron bandas definidas y, luego de la secuenciación, se presentaron negativas para Hepacivirus. En análisis evolutivos se observó que el virus GB-C ocupa la posición filogenético más distante en relación al VHC, mientras que la más próxima es ocupada por el virus GB-B. A pesar de la no identificación de Hepacivirus parecidos al VHC en primates no humanos, no se puede afirmar que el mismo no exista. Estudios involucrando a otras especies, como los virus GBs y los posibles nuevos Hepacivirus pueden solucionar el proceso evolutivo que permitió que el VHC infectara a seres humanos, ya que estos parecen ser el eslabón entre el VHC y las especies virales encontradas en animales silvestres.
Palabras clave: Hepacivirus; Pegivirus; Virus GB-A; Virus GB-B; Virus GB-C.
INTRODUÇÃO
A classificação e a relação filogenética viral são definidas a partir de vários fatores como morfologia, biologia, natureza e sequência do ácido nucleico. Nesse processo, as ferramentas da biologia molecular vêm sendo utilizadas de forma decisiva, devido sua grande sensibilidade e especificidade. Assim, diversas espécies virais foram e continuam sendo identificadas a cada ano.
A partir da identificação de novas espécies virais é possível esclarecer inúmeros processos coevolutivos, biogeográficos, biológicos e epidemiológicos de importância. Pela proximidade evolutiva envolvendo os primatas não humanos e humanos, existe maior probabilidade de encontrarem-se vírus filogeneticamente relacionados envolvendo tais reservatórios.
A família Hepadnaviridae é um exemplo típico da relação evolutiva envolvendo vírus e espécies hospedeiras, havendo claramente uma subdivisão em dois grupos de vertebrados. Assim, aves são infectadas por espécies virais que constituem o gênero Avehepadnavirus e mamíferos por aquelas que constituem o gênero Orthohepadnavirus1,2,3. Dentre os que afetam mamíferos existem aqueles mais proximamente relacionados aos roedores (vírus da hepatite B - VHB da marmota) e aqueles mais proximamente relacionados aos primatas não humanos (VHB de primatas não humanos) e humanos (VHB).
Na família Flaviviridae são encontradas também relações coevolutivas claras, em particular para o gênero Hepacivirus, formado pelo vírus da hepatite C (VHC). Alguns autores classificam também no gênero os vírus GB A, vírus GB B, vírus GB C, vírus GB D e o VHC, sendo as espécies do gênero Saguinus (Primates, Platyrrhini), reservatórios de vírus GB A e vírus GB B, enquanto o homem é reservatório de vírus GB C e do VHC. Estes últimos apresentam como diferença biológica um tropismo tecidual particular, linfotrópico e hepatotrópico, respectivamente. No entanto, do ponto de vista molecular, vírus GB A e vírus GB C apresentam semelhanças moleculares na região 5' não transcrita (5' UTR) com VHC, sugerindo uma biologia e, portanto, epidemiologia, em comum4,5,6,7,8.
Stapleton et al9 propõem a classificação dos vírus GB A, vírus GB C e vírus GB D em um novo gênero, Pegivirus, dentro de Flaviviridae, baseando principalmente na ideia de que estes vírus não causam hepatite em seus hospedeiros. O vírus GB A e o vírus GB C seriam chamados de pegivírus A, sendo uma única espécie com hospedeiros diferentes; e o vírus GB D seria chamado de pegivírus B. Recentemente o Comitê Internacional de Taxonomia Viral (International Committee on Taxonomy of Viruses - ICTV) aceitou tais classificações10.
Os recentemente encontrados vírus semelhantes a hepacivírus, chamados de hepacivírus de cães (CHV, canine hepacivirus), hepacivírus de cavalos (NPHV) e hepacivírus de roedores (rodent hepacivirus) ainda não possuem uma classificação certa. Pela grande semelhança genômica descrita por Kapoor et al11 para o hepacivírus canino, foi proposta sua classificação no gênero Hepacivirus, o mesmo ocorrendo na descrição do hepacivírus de roedores (Myodes giareoius e Rhabdomys pumilio) por Drexler et al12. Entretanto, nenhuma destas classificações de novos vírus foram aceitas pelo ICTV até o presente momento10. Vários autores acreditam que o vírus GB B deve ser classificado junto com o VHC em Hepacivirus graças às suas similaridades genômicas e às suas epidemiologia e hepatotropia9.
O VHC apresenta atualmente uma grande importância epidemiológica por que responde por uma grande epidemia mundial, com aproximadamente 180 milhões de indivíduos infectados13. Desde a sua descoberta14, esse vírus vem sendo estudado a fim de se ter sua melhor caracterização. Assim, Kreukulová et al15 caracterizaram molecularmente o VHC, identificando um genoma de RNA positivo formado por cerca de 9.600 nucleotídeos com duas regiões não codificantes, nas posições 5' e 3', respectivamente, e uma região intermediária codificante de uma poliproteína de 3.010 a 3.033 aminoácidos, a qual é clivada, formando proteínas estruturais (C, p7, E1 e E2) e não estruturais (NS2, NS3, NS4A, NS4B, NS5A e NS5B).
Na extremidade 5' da sequência nucleotídica do VHC há uma região altamente conservada, a internal ribosome entry site (IRES), formada por uma sequência de 315 nucleotídeos, fundamental para a ligação da fita de RNA viral com o ribossomo hospedeiro e tradução da mesma, portanto necessária para a replicação viral15,16,17. Sabe-se que a região IRES promove formação correta do complexo ribossômico 48S com a necessidade somente dos fatores de iniciação eIF3 e do complexo eIF2+Met-tRNAiMet+GTP e mudando sua conformação no contato com o rRNA 18S para a ligação do rRNA 40S18.
A conservação da IRES se deve à grande pressão seletiva a ela relacionada, devido à sua importância na replicação viral. Por esse motivo essa região foi escolhida como referência para a identificação de outras espécies virais semelhantes ao VHC em primatas não humanos.
Apesar do conhecimento relativamente reduzido em relação à evolução viral, particularmente dos vírus de RNA, existem evidências de que grupos virais com ecologias particulares apresentam uma taxa de evolução reduzida19. No entanto, a maioria dos dados aponta para uma grande capacidade evolutiva dos vírus de RNA, se comparados aos vírus de DNA, o que ocorre devida a algumas características particulares, como alta taxa de mutação, genoma de tamanho reduzido, grande capacidade de adaptação, fortes efeitos deletérios provocados pela deriva genética e elevada sensibilidade ao acúmulo de mutações20.
Devido a todas as características evolutivas dos vírus de RNA, esses estariam sujeitos a diferentes leis evolutivas21, o que tem sido confirmado por estudos epidemiológicos, funcionais e estruturais, os quais indicam que os vírus de RNA podem tolerar tipos e números restritos de mutação durante a sua evolução.
A despeito de sua grande capacidade evolutiva, os vírus de RNA são capazes de associações estáveis com seus hospedeiros, ao longo dos processos coevolutivos22. Essa capacidade evolutiva tornou-se evidente a partir das duas últimas décadas, após a constatação de que os vírus de RNA possuem a capacidade de combinar o seu material genético com os de outros vírus, assim como adquirir genes de seus hospedeiros, eliminar mutações deletérias acumuladas e dispersar combinações benéficas de mutações, realizando, nesse processo, recombinações genéticas, as quais geram sequencias nucleotídicas com diferenças significativas daquelas originais. Tal processo ocorre em espécies virais de RNA segmentado ou não segmentado23. Desse modo, os vírus de RNA apresentam uma grande capacidade de se adaptar a novos ambientes, novas pressões seletivas e novos hospedeiros, quando surgem as oportunidades24.
Até o presente momento não existem informações sobre quais mutações sofridas pelo predecessor do VHC o tornaram capaz de infectar seres humanos por que tal predecessor ainda não foi encontrado. Os vírus GBs parecem ter uma relação mais próxima com a hipotética espécie viral, já que são encontrados em primatas não humanos e em humanos. No entanto, de acordo com Domingo et al25, os vírus GBs podem ter surgido a partirde mutações nos genomas de outros vírus, como o VHC ou o VHB, indicando estas espécies virais como as mais primitivas.
As recentes descobertas do NPHV, CHV e do hepacivírus de roedores, possuindo semelhanças com o VHC, talvez quebre esse pressuposto de que o parente mais próximo do VHC seria encontrado em primatas não humanos. Entretanto, a dúvida ainda permanece, pois tais semelhanças são insuficientes para propor uma origem do VHC. Acredita-se que ainda podemos encontrar Hepacivirus em outras espécies de mamíferos, principalmente primatas não humanos, como ocorreu com HIV-1, do qual sabe-se que teve origem em chimpanzés26.
Entretanto, Simmonds26 também cita a teoria de que o VHC pode não ter uma origem a partir de animais silvestres, sendo apenas um vírus que sempre infectou seres humanos, hominídeos e provavelmente o ancestral dos mamíferos.
A descoberta de uma origem e um possível hospedeiro primário que pode ter levado a várias epidemias causadas pela infecção pelo VHC, varrendo a população humana em séculos passados, seria um passo importante na compreensão das relações com o ser humano, sua adaptação e patogenicidade26.
Dessa maneira, o objetivo deste trabalho é pesquisar a presença de Hepacivirus semelhantes ao VHC, em amostras de soros de primatas não humanos, do Velho e do Novo Mundo, mantidos no criobanco da Seção de Hepatologia (SAHEP) do Instituto Evandro Chagas (IEC).
MATERIAIS E MÉTODOS
DELIMITAÇÃO DO ESTUDO
Todos os procedimentos para este trabalho foram realizados no IEC. A pesquisa foi aprovada pelo Comitê de Ética em Pesquisas com Animais do IEC, sob o número de registro 0029/2008/IEC/SVS/MS.
Os ácidos nucleicos virais, RNAs, foram obtidos de 1.054 amostras de soros de primatas não humanos depositadas no criobanco da SAHEP/IEC, pertencentes a 34 espécies de primatas não humanos do novo mundo: Alouatta belzebul, Alouatta caraya, Alouatta seniculus, Aotus azarae, Aotus infulatus, Aotus trivigartus, Ateles belzebuth marginatus, Ateles paniscus, Callimico goeldii, Callicebus molock, Callicebus torquatus, Callithrix emiliae, Callithrix geoffroyi, Callithrix jacchus, Callithrix penicilata, Cebus albifrons, Cebus apella, Cebus nigrivitatus, Cebus olivaceus, Cebus xanthosthernus, Cebuella pygmea, Chiropotes satanas, Lagothrix lagothricha, Pithecia irroratta, Pithecia pithecia, Saimiri boliviensis, Saimiri sciureus sciureus, Saimiri ustus, Saguinus fuscicollis weddelli, Saguinus imperator, Saguinus labiatus, Saguinus midas e Saguinus niger. E duas espécies de primatas não humanos do velho mundo: Cercopithecus aethiops e Pan troglodytes.
INICIADORES
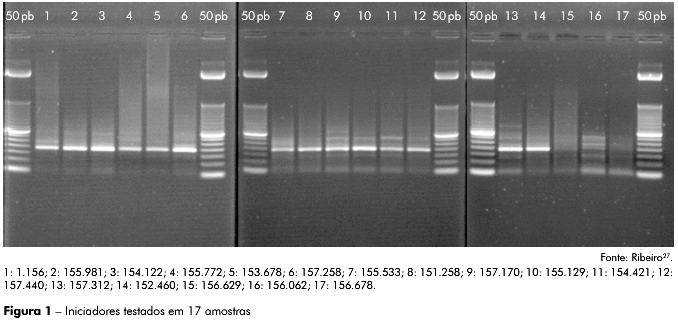
Foram utilizados iniciadores desenhados e testados na SAHEP/IEC para o diagnóstico de Hepacivirus semelhantes ao VHC, indicando especificidade unicamente para o VHC e negatividade para os vírus da hepatite A (VHA), VHB e vírus da hepatite D (VHD). A sensibilidade dos iniciadores foi de 850.000 UI/mL a menos que 600 UI/mL27 (Figura 1): APE1 (CCC TGC GCG GYA AYA RGY A), APE2 (GGG AGA GCC TTR GTG GTC TG) e APE3 (GGG CAC TYG CAA GCA CCS T), os quais se ligam nas regiões 449-467, 133-153 e 302-321, respectivamente, da molécula de RNA do VHC, espécie do gênero Hepacivirus. Essa região compreende parte da IRES, presente na região 5' UTR, sendo o produto de amplificação final um amplicon de 188 pares de nucleotídeos.
PROCEDIMENTOS COM A TÉCNICA DE PCR
A extração de RNA foi feita pelo método com TRIzol® LS Reagent, Invitrogen28, de acordo com o fabricante.
Para obtenção de moléculas de DNA complementares, os RNAs obtidos do soro de primatas não humanos foram submetidos à técnica de RT-PCR, a partir do protocolo: 5X First-Stand Buffer (4 µL), DTT-0,1 M (2 µL), DNTPs -10 mM (1 µL), RNAsin-10 U (1 µL), iniciador APE1 (1 µL), M-MLV-200 U (1 µL), suspensão de RNA (10 µL). Para as temperaturas: 37o C por 1 h, 95o C por 15 min e um Hold de 4o C.
A fim de obter uma maior quantidade de amplicons para a detecção em gel de agarose, procedeu-se à técnica de PCR para o diagnóstico de Hepacivirus, a partir do protocolo: água destilada e deionizada (30 µL), 10X PCR Buffer (5 µL), Cloreto de Magnésio (1,5 µL), DNTPs-10 mM (1 µl), iniciadores APE2 e APE3 (1 µL cada), TAQ-2,5 U (0,5 µL), suspensão de DNA complementar (10 µL). Para as temperaturas: 94o C por 1 min, 35 ciclos de 94o C por 1 min, 55o C por 1 min, 72o C por 1 min e 30 seg, 72o C por 7 min e Hold de 4o C. Para a obtenção de uma quantidade de amplicons detectáveis por eletroforese, foram realizadas duas PCRs para cada amostra.
Adicionalmente, foram utilizados controles positivos, constituídos de amostras sorológicas positivas para o VHC, presentes na soroteca da SAHEP/IEC, identificados como CP-153.678 e CP-169.666 (controles positivos).
SEQUENCIAMENTO
A quantificação foi realizada em gel de Agarose a 2%, aplicando 4 µL das amostras, 2 µL de Ladder e 4 µL de Low Mass Ladder. E para o sequenciamento foi utilizado o kit Big-Dye Terminator (Applied Biosystems) e o protocolo segundo o seu fabricante, utilizando o sequenciador ABI PRISM 3130 XL (Applied Biosystems).
COMPARAÇÃO DE SEQUÊNCIAS NUCLEOTÍDICAS
As sequências nucleotídicas obtidas por sequenciamento e pelo Genbank foram analisadas utilizando a ferramenta online BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi) e comparadas utilizando o BioEdit 7.0.
RESULTADOS
Das 1.054 amostras analisadas obtivemos como resultados: 1.041 sem amplificação, 11 apresentando uma banda inespecífica (menor ou maior que 188 pb, de tamanho variando entre 60 pb e 250 pb), as quais foram mantidas, por serem prováveis espécies virais filogeneticamente relacionadas, e uma amostra com banda específica para 188 pb, como apresentado na figura 2, figura 3 e tabela 1.
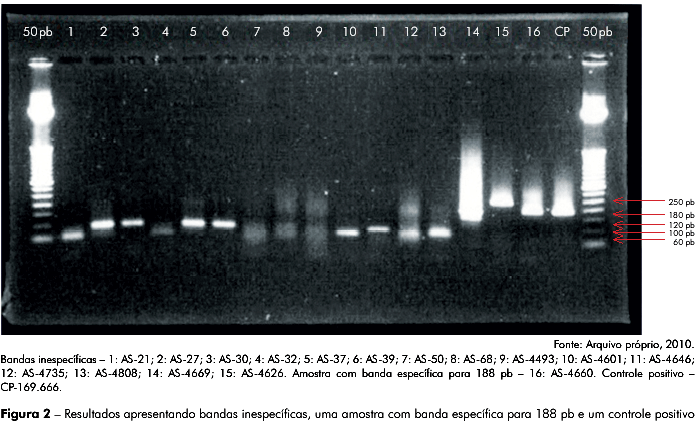
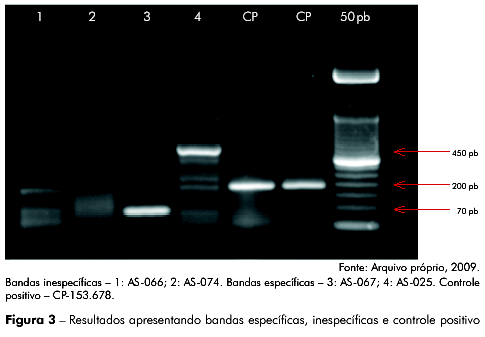

Todos os amplicons com bandas inespecíficas únicas e específicas para 188 pb, assim como dos controles positivos CP-153.678 e CP-169.666 foram sequenciados, apresentando resultado negativo para a região IRES do VHC, nas amostras sorológicas de primatas não humanos, e resultado positivo para os controles positivos utilizados (CP-153.678 e CP-169.666), como esperado, comprovando a especificidade e a sensibilidade dos iniciadores desenhados para este estudo (Figura 1).
DISCUSSÃO
Semelhante aos resultados desta pesquisa, Makuwa et al29 realizaram buscas de hepatites virais em espécies selvagens de primatas não humanos encontradas no País africano Gabão, não encontrando positividade para o VHC, somente para o VHB e VHD. Porém, a diferença entre os dois trabalhos é que, neste, utilizaram-se marcadores moleculares de biologia molecular, enquanto Makuwa et al29 basearam-se principalmente em marcadores sorológicos.
Em análises realizadas previamente6 identificou-se uma maior proximidade filogenética entre os vírus GB B e o VHC, do que com as outras espécies de vírus GBs (vírus GB A/pegivírus A e vírus GB C/pegivírus A), identificando-se uma maior semelhança envolvendo sequências nucleotídicas conservadas do gene NS3 (Figura 4), o qual codifica as proteínas não estruturais: proteinase, ATPase e helicase. Outra fonte de semelhança é a região de clivagem entre NS4A e NS4B e entre NS4B e NS5A. Dessa maneira, para Hepacivirus, todas essas sequências parecem apresentar uma maior importância no processo de replicação viral do que a própria IRES, presente em Hepacivirus.
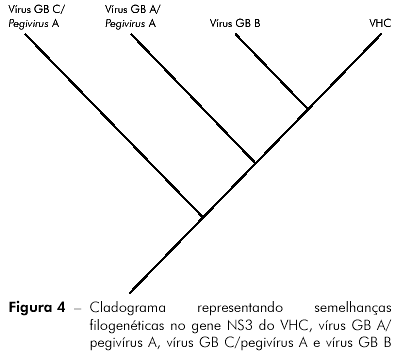
Com a futura identificação de correspondentes virais filogeneticamente relacionados ao VHC em primatas não humanos e a utilização de um grupo externo adequado para as polarizações da árvore filogenética, poderemos identificar, ao longo da evolução de Hepacivirus, se houve um acréscimo ou uma perda da IRES, o que também indicará o sentido do cruzamento da barreira evolutiva entre as espécies reservatórias.
Do ponto de vista filogenético, parece improvável a existência de um gênero viral com poucas espécies, o que foi confirmado pela identificação recente de uma nova espécie viral, denominado GBV-D, o qual, na comparação com os demais Hepacivirus, apresentou grande semelhança com vírus GB A e vírus GB C para os genes NS3 e NS5. No entanto, devido a características próprias desse vírus, os descobridores dele sugeriram que ele seja classificado em outro gênero de Flaviviridae4, o que foi aceito recentemente pelo ICTV no novo gênero Pegivirus. Sendo assim, os genes NS3 e NS5, do GBV-D parecem de grande importância na identificação do elo filogenético de ligação entre VHC e possíveis Hepacivirus presentes em primatas não humanos.
Reforçando a teoria da existência de outros Hepacivirus ainda não classificados, mais recentemente foram encontrados o CHV, o hepacivírus não primata de cavalos (NPHV-posifive horses) e o hepacivirus de roedores30, os quais necessitam de análises moleculares mais completas para elucidar sua relação filogenética com os Hepacivirus já identificados.
De acordo com o trabalho de Drexler et al12, os hepacivírus de roedores seriam um grupo irmão do vírus GB B, e o VHC, um grupo irmão do CHV, e por isso ele propõe que o vírus GB Beos hepacivirus de roedores devem ser classificados em um novo gênero dentro de Flaviviridae, por apresentarem status monofilético em suas análises, enquanto o CHV, por possuir grande semelhança genética com o VHC, deve ser classificado em Hepacivirus. Em contraste, no trabalho de Stapleton et al9, os autores propõem a classificação do vírus GB B e o do hepacivírus de roedores em Hepacivirus.
Lyons et al30 e Kapoor et al11 descreveram grande semelhança genética entre o VHC e o CHV e propõem a sua classificação em Hepacivirus. Porém, estes novos Hepacivirus, incluindo o NPHV, devem ser melhor estudados, desde seu genoma, epidemiologia e estrutura morfológica para que sejam realmente classificados no gênero Hepacivirus, principalmente por ter o CHV sido encontrado em maior quantidade nos pulmões do que no fígado, diferenciando-se do VHC, que é hepatotrópico.
Simmonds26 cita a teoria de que esses novos possíveis Hepacivirus podem ter surgido de outros vírus ainda não encontrados ou do próprio VHC. Uma evidência disto pode se dever ao NPHV possuir a protease responsável pela clivagem da MAVS (proteína mitocondrial de sinalização antiviral) e TRIF (domínio TIR adaptador de indução do IFN-P) humano, prevenindo, assim a resposta antiviral da célula e potencializando a replicação viral. Outra teoria é a de que cada espécie de Hepacivirus, ou possíveis Hepacivirus, se adapta exclusivamente ao seu hospedeiro, como o VHC em humanos, o NPHV em cavalos, o CHV em roedores26.
CONCLUSÃO
Devido à grande capacidade de mutação e recombinação genética presente nos vírus de RNA e a negatividade em nossos resultados para um vírus com a IRES semelhante a do VHC em primatas não humanos, conclui-se que, provavelmente, o grupo com IRES presente em animais silvestres teria sido eliminado pela seleção natural, após mutações deletérias, ou teria ocorrido uma recombinação genética, envolvendo moléculas de RNA virais, conferindo a IRES ao VHC, indicando uma possível recombinação molecular.
Como não se verificou a presença de Hepacivirus nas amostras sorológicas de primatas não humanos do Novo e Velho Mundo, para a sequência altamente conservada do VHC, a sequência 5' UTR - IRES, infere-se que os vírus GBs, encontrados em animais silvestres e novos possíveis Hepacivirus, podem ser mais proximamente relacionados ao VHC. Sendo assim, os próximos estudos com objetivo de encontrar correspondentes virais ao VHC em primatas não humanos, devem concentrar-se em outras sequências nucleotídicas compartilhadas diferentes da IRES, já que, utilizando-se a mesma, não foi encontrada positividade.
A negatividade de Hepacivirus, utilizando a região IRES, em primatas não humanos e a recente descoberta de novos vírus em outros mamíferos não exclui a possibilidade de origem do VHC em primatas não humanos, pois, além de análises complementares de epidemiologia e morfologia, buscas em uma variedade maior de mamíferos, incluindo espécies de primatas não humanos, ainda devem ser feita para finalmente propor-se uma origem para o VHC e para elucidar a verdadeira relação filogenética com os novos possíveis Hepacivirus.
AGRADECIMENTOS
Ao dr. Pedro Vasconcelos, chefe da Seção de Arbovirologia e Febres Hemorrágicas do IEC. Aos amigos e profissionais das seções de Virologia, Arbovirologia e Febres Hemorrágicas, Hepatologia e do Núcleo de Análise Genética por Imagem.
REFERÊNCIAS
1 Knipe DM, Howley PM, editors. Fields virology. 5th ed. Philadelphia: Lippincott Williams and Wilkings; 2006. 2 v.
2 Funk A, Mhamdi M, Willi H, Sirma H. Avian hepatitis B viruses: molecular and cellular biology, phylogenesis, and host tropism. World J Gastroenterol. 2007 Jan;13(1):91-103. [Link]
3 Glebe D, Urban S. Viral and cellular determinants involved in hepadnaviral entry. World J Gastroenterol. 2007 Jan;13(1):22-38. [Link]
4 Epstein JH, Quan PL, Briese T, Street C, Jabado O, Conlan S, et al. Identification of GBV-D, a novel GBV-like flavivirus from old world frugivorous bats (Pteropus giganteus) in Bangladesh. PLoS Pathog. 2010 Jul;6(7):e1000972. Doi:10.1371/journal.ppat.1000972 [Link]
5 Simmonds P. The origin and evolution of hepatitis viruses in humans. J Gen Virol. 2001 Apr;82(Pt 4):693-712. [Link]
6 Beames B, Chavez D, Lanford RE. GB virus as a model for hepatitis C virus. ILAR J. 2001;42(2):152-60. Doi:10.1093/ilar.42.2.152 [Link]
7 Mansfield KG. GB virus B model of hepatitis C infection. In: 55th Annual Meeting of the American College of Veterinary Pathologists and 39th Annual Meeting of the American Society of Clinical Pathology; 2004 Nov 13; Middleton: American College of Veterinary Pathologists and American Society for Veterinary Clinical Pathology; 2004. [Link]
8 Lindenback BD, Rice CM. Flaviviridae: the viruses and their replications. In: Knipe DM, Howley PM, Griffin DE, editors. Fields virology. 4th ed. Philadelphia: Lippincoot Williams & Wilkins; 2001. p. 991-1041.
9 Stapleton JT, Foung S, Muerhoff AS, Bukh J, Simmonds P. The GB viruses: a review and proposed classification of GBV-A, GBV-C (HGV), and GBV-D in genus Pegivirus within the family Flaviviridae. J Gen Virol. 2011 Feb;91(Pt):233-46. Doi:10.1099/vir.0.027490-0 [Link]
10 International Committee on Taxonomy of Viruses [Internet]. 2013 Set [cited 2013 Set] Available from: http://www.ictvonline.org/virusTaxonomy.asp.
11 Kapoor A, Simmonds P, Gerold G, Qaisar N, Jain K, Henriquez JA, et al. Characterization of canine homolog of hepatites C virus. Proc Natl Acad Sci U S A. 2011 Jul;108(28):11608-13. Doi: 10.1073/pnas.1101794108 [Link]
12 Drexler JF, Corman VM, Müller MA, Lukashev AN, Gmyl A, Coutard B, et al. Evidence for novel Hepaciviruses in rodents. PLoS Pathogens. 2013;9(6):1-17. Doi:10.1371/journal.ppat.1003438 [Link]
13 Ciorlia LAS, Zanetta DMT. Hepatite C em profissionais da saúde: prevalência e associação com fatores de risco. Rev Saude Publica. 2007 Apr;41(2):229-35. Doi:10.1590/S0034-89102007000200009 [Link]
14 Choo QL, Kuo G, Weiner AJ, Overby LR, Bradley DW, Houghton M. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science. 1989 Apr;244(4902):359-62. Doi:10.1126/science.2523562 [Link]
15 Krekulová L, Rehák V, Riley LW. Structure and functions of hepatitis C virus proteins: 15 years after. Folia Microbiol. 2006;51(6):665-80. [Link]
16 Brass V, Moradpour D, Blum HE. Molecular virology of hepatitis C virus (HCV): 2006 Update. Int J Med Sci. 2006;3(2):29-34. Doi:10.7150/ijms.3.29 [Link]
17 Helen CUT, Breyne S. A distinct group of hepacivirus/pestivirus-like internal ribosomal entry sites in members of diverse Picornavirus genera: evidence for modular exchange of functional noncoding RNA elements by recombination. J Virol. 2007 Jun;81(11):5850-63. Doi:10.1128/JVI.02403-06 [Link]
18 Malygin AA, Kossinova OA, Shatsky IN, Karpova GG. HCV IRES interacts with the 18S rRNA to activate the 40S ribosome for subsequent steps of translation initiation. Nucleic Acids Res. 2013 Jul:1-9. Doi:10.1093/nar/gkt632 [Link]
19 Jenkins GM, Rambaut A, Pybus OG, Holmes EC. Rates of molecular evolution in RNA viruses: a quantitative phylogenetic analysis. J Mol Evol. 2002 Feb;54(2):156-65. [Link]
20 Cuevas JM, Domingo-Calap P, Pereira-Gómez M, Sanjuán R. Experimental evolution and population genetics of RNA viruses. The Op Evol J. 2009;3:9-16. Doi:10.2174/1874404400903010009 [Link]
21 Holmes EC, Moya A. Is the quasispecies concept relevant to RNA viruses? J Virol. 2002 Jan;76(1):460-2. Doi:10.1128/JVI.76.1.460-462.2002 [Link]
22 Jackson AP, Charleston MA. A cophylogenetic perspective of RNA-virus evolution. Mol Biol Evol. 2004 Jan;21(1):45-57. Doi:10.1093/molbev/msg232 [Link]
23 Worobey M, Holmes EC. Evolutionary aspects of recombination in RNA viruses. J Gen Virol. 1999 Oct;80(Pt 10):2535-43. [Link]
24 Simmonds P. RNA viruses: evolution in action. Microbiol Today. 2004;31:163-5. [Link]
25 Domingo E, Gomez J. Quasispecies and its impact on viral hepatitis. Virus Res. 2007 Aug;127(2):131-50. Doi:10.1016/j.virusres.2007.02.001 [Link]
26 Simmonds P. The origin of hepatitis C virus. Curr Top Microbiol Immunol. 2013;369:1-15. Doi:10.1007/978-3-642-27340-7_1 [Link]
27 Ribeiro NAB. Documento técnico apresentando desenho de iniciadores de espécies do gênero Hepacivirus, baseado no alinhamento de sequencias nucleotídicas disponíveis. UNESCO; 2008.
28 Chomczynski P, Sacchi N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem. 1987 Apr;162(1):156-9. Doi:10.1016/0003-2697(87)90021-2 [Link]
29 Makuwa M, Souquiére S, Telfer P, Bourry O, Rouquet P, Kazanji M, et al. Hepatitis viruses in non-human primates. J Med Primatol. 2006 Dec; 35(6):384-7. Doi:10.1111/j.1600-0684.2006.00163.x [Link]
30 Lyons S, Kapoor A, Sharp C, Schneider BS, Wolfe ND, Culshaw G, et al. Nonprimate hepaciviruses in domestic horses, United Kingdom. Emerg Infect Dis. 2012 Dec;18(12):1976-82. Doi:10.3201/eid1812.120498 [Link]
 Correspondência / Correspondence / Correspondencia:
Correspondência / Correspondence / Correspondencia:
Gustavo Moraes Holanda
Rua Antonio Barreto, 747. Bairro: Umarizal
CEP:66055-050 Belém-Pará-Brasil
Tel.: +55 (91) 8807-4903
E-mail: holandagm@gmail.com
Recebido em / Received / Recibido en: 11/3/2013
Aceito em / Accepted / Aceito en: 24/9/2013
