Caracterização morfoanatômica dos caules de Cyperus articulatus L. e C. prolixus H.B.K. (Cyperaceae)
Morpho-anatomical characterization of stems of Cyperus articulatus L. and C. prolixus H.B.K. (Cyperaceae)
Pedro Paulo dos SantosI, Raimunda Conceição de Vilhena PotiguaraI, II, Alba Lúcia Ferreira de Almeida LinsI, Eunice Gonçalves MacedoIII
IMuseu Paraense Emílio Goeldi/Ministério da Ciência, Tecnologia e Inovação. Belém, Pará, Brasil
IIIn memorian
IIIUniversidade do Estado do Pará. Belém, Pará, Brasil
RESUMO
Cyperus articulatus e C. prolixus são conhecidas, respectivamente, como priprioca e pripriocão. Nessas espécies, há discordâncias quanto à nomenclatura de seus caules, que recebem denominações diversas, provocando dificuldades na identificação dos taxa. O presente estudo objetivou caracterizar os caules das duas espécies por meio da morfologia, morfometria, funcionalidade, anatomia e histoquímica. Os espécimes foram coletados em experimentos do campus Belém da Universidade Federal Rural da Amazônia (UFRA), sendo sua procedência do município de Acará, estado do Pará. O material foi descrito, mensurado e fixado em FAA70, seguindo-se, posteriormente, as práticas usuais em anatomia vegetal e testes histoquímicos. Os caules são dos tipos rizoma e cormo, subterrâneos, ambos plagiotrópicos e revestidos por catafilos. O rizoma é monopodial e cilíndrico, já o cormo é intumescido. Os dois caules são perenes; o cormo possui a função de propagação e o rizoma de expansão caulinar. Quanto à anatomia, os caules apresentam meristema de espessamento primário ativo, porém, apenas os cormos possuem meristema de espessamento secundário e produzem raízes adventícias.
Palavras-chave: Monocotiledônea. Rizoma. Cormo.
ABSTRACT
Cyperus articulatus and C. prolixus are known as priprioca and pripriocão, respectively. In these species, there are disagreements on the nomenclature of the stems, which receive different names, causing difficulties in the taxa identification. The objective of this paper is the characterization of the C. articulatus and C. prolixus stems, through the description of morphology, morphometric data, function, anatomy and histochemistry. Samples were collected in a growth experiment in the Universidade Federal Rural da Amazônia (UFRA), Belém municipality, and the plants are originally from Acará municipality, Pará state. The material was described, measured, fixed in FAA70, and studied following usual anatomy techniques and histochemical tests. The stems are of two types rhizome and corm. They are subterranean, plagiotropic and surrounded by cataphylls. The rhizome is monopodial and cylindrical, while the corm is swollen. These two stems are perennial, the corm has propagation function and the rhizome has expansion function. Regarding anatomy, these stems have active primary thickening meristem, but only corms have secondary thickening meristem activity and produce adventitious roots.
Keywords: Monocotyledoneae. Rhizome. Corm.
]]>
INTRODUÇÃO
O território brasileiro possui cerca de 680 espécies de Cyperaceae, correspondendo a mais de 15% do total de representantes dessa família ocorrentes no mundo (Alves et al., 2009). São geralmente ervas perenes, rizomatosas, cosmopolitas, de habitat úmido, com corpo vegetativo constituído por raízes, folhas e escapo triangular em secção transversal (Metcalfe, 1971; Judd et al., 2009).
Entre os representantes da flora de Cyperaceae brasileira estão Cyperus articulatus e C. prolixus. Elas são conhecidas, segundo Zoghbi et al. (2008), como priprioca e pripriocão. Além disso, são integrantes do secular e tradicional banho perfumado chamado cheiro-do-pará.
Potiguara & Santos (2008) relataram que em tais espécies há discordância quanto à nomenclatura dos constituintes do sistema subterrâneo, e que esta porção vegetativa recebe denominações de raízes tuberosas, tubérculos ou rizoma.
Recentes estudos realizados no Brasil, entre esses, os de Estelita & Rodrigues (2007) e Rodrigues & Estelita (2009), estão reinterpretando a origem e nomenclatura dos órgãos subterrâneos e aéreos das Cyperaceae, a partir de características anatômicas, como a atuação dos meristemas de espessamento primário e secundário, do período de vida e da função.
Nessa nova perspectiva de caracterização morfológica, percebeu-se que o sistema subterrâneo de Cyperaceae pode apresentar comportamentos adaptativos complexos, culminando com mais de um tipo de caule no mesmo indivíduo.
Este estudo objetivou caracterizar os caules de Cyperus articulatus e C. prolixus por meio de dados morfológicos, morfométricos, funcionais, anatômicos e histoquímicos.
MATERIAL E MÉTODOS
]]> Foram analisadas amostras de Cyperus articulatus e de C. prolixus coletadas de experimentos da Universidade Federal Rural da Amazônia, sendo as plantas procedentes da comunidade de Boa Vista, Acará, Pará. Duas amostras foram depositadas no herbário João Murça Pires, do Museu Paraense Emílio Goeldi (MG), com os respectivos números de registros: C. articulatus - MG 167.227 e C. prolixus - MG 178.403.Para a análise morfológica, foi utilizada a nomenclatura de Gonçalves & Lorenzi (2007) para tipos de caules subterrâneos.
As descrições morfológicas e os dados morfométricos foram realizados em 20 amostras de cada espécie, sendo mensurados o comprimento e a largura dos órgãos caulinares com o auxílio de paquímetro e trena.
Para a análise anatômica, os caules foram fragmentados e fixados em FAA70, por 24 horas, lavados e conservados em etanol 70% (Johansen, 1940).
Os fragmentos das amostras do caule localizados até o quinto nó de afastamento do ápice caulinar foram tratados para infiltração e inclusão em parafina (Johansen, 1940). Logo após, esse material foi seccionado com auxílio de micrótomo rotativo, tendo sido corado com azul de Astra 1% e fucsina básica 1% (Braga, 1977). Posteriormente, as lâminas foram montadas em bálsamo do Canadá.
Para os testes histoquímicos, foram realizadas secções à mão livre com o auxílio de lâmina de barbear. Foi utilizado lugol para determinação de amido; floroglucinol com ácido clorídrico 1% para diagnóstico de lignina; cloreto férrico 10% para compostos fenólicos; e sudam iii para óleos e suberina (Johansen, 1940). Testes controle foram realizados conforme recomendação de Johansen (1940).
As fotomicrografias foram obtidas com câmera digital acoplada ao microscópio óptico e os desenhos à mão livre, em microscópio estereoscópio.
RESULTADOS
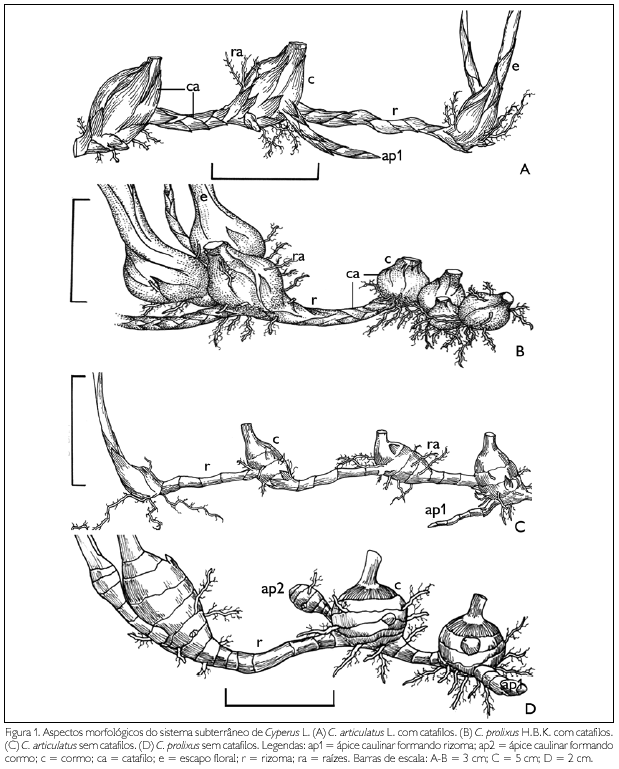
Os caules de Cyperus articulatus e C. prolixus são subterrâneos, apresentando crescimento plagiotrópico. São monopodiais e estão envolvidos por catafilos (Figuras 1A-1D). Os caules jovens possuem coloração esbranquiçada e os adultos apresentam coloração escura.
]]>
Em ambas as espécies, o caule possui uma porção cilíndrica, o rizoma, e outra intumescida, o cormo (Figuras 1A-1D), que derivam do ápice caulinar vegetativo, logo após os primeiros nós (Figura 1D).
O cormo é perene, varia da forma elíptica a esférica, de consistência rígida, localizando-se entre os rizomas (Figuras 1A-1D). Apresenta comprimento de 0,8 a 2,5 cm e largura de 0,5 a 2 cm, em C. articulatus, e comprimento 1,3 a 3 cm e largura de 1 a 2 cm em C. prolixus, sendo a primeira espécie com dois a quatro nós, e a segunda com três a sete nós. Os catafilos que os envolvem possuem forma linear, de um a três por nó, em filotaxia verticilata, paralelinérveas, comprimento 0,6 a 0,9 cm e largura 0,3 a 0,5 cm, em C. articulatus, e comprimento 0,6 a 1,6 cm e largura 0,5 a 0,7 cm em C. prolixus.
A partir do cormo, origina-se o escapo floral para a superfície, o ápice caulinar ou botões apicais na propagação vegetativa (Figuras 1A-1D) e as raízes exclusivamente adventícias de nós e entrenós (Figuras 1C e 1D), o que não ocorre no rizoma.
O rizoma é perene, cilíndrico, delgado e alonga-se no sentido horizontal, de consistência esponjosa em C. articulatus e rígido em C. prolixus. Apresenta comprimento de 1 a 6 cm e largura de 0,3 a 0,5 cm, em C. articulatus, comprimento de 0,4 a 3,5 cm e largura de 0,6 a 2 cm em C. prolixus, possuindo três a oito nós, na primeira espécie, e dois a sete na segunda (Figuras 1C e 1D). Os catafilos dos rizomas diferem dos encontrados no cormo apenas nas dimensões, sendo o comprimento de 0,6 a 0,9 cm e largura de 0,3 a 0,4 cm, em C. articulatus, e comprimento de 0,5 a 1 cm e largura de 0,3 a 0,4 cm em C. prolixus. Nesta última espécie, os cormos podem ocorrer próximos, interrompidos por rizomas (Figura 1B) de, no mínimo, dois nós.
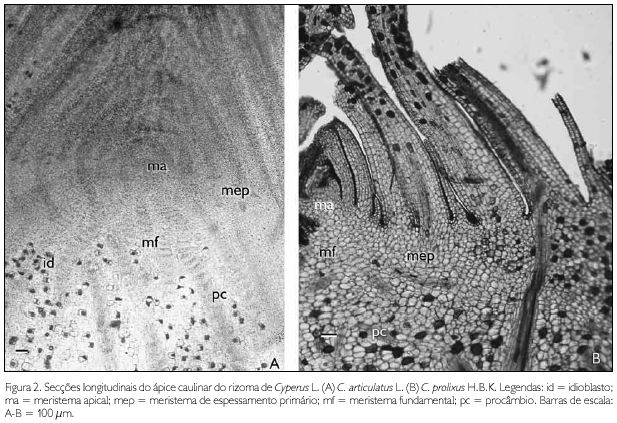
Anatomicamente, nas duas espécies, os ápices caulinares do cormo e do rizoma apresentam meristema apical, meristema fundamental, procâmbio e meristema de espessamento primário (MEP) ativos (Figuras 2A e 2B). O MEP é reconhecido por camadas de células de parede delgadas, em constante divisão, envolvendo o meristema fundamental e o procâmbio (Figura 2B). As células derivadas do MEP formam parênquima e feixes vasculares, delimitando o córtex e a medula no cormo e no rizoma (Figuras 3A e 3B). No cormo, este meristema também forma raízes adventícias (Figura 3C).
]]>
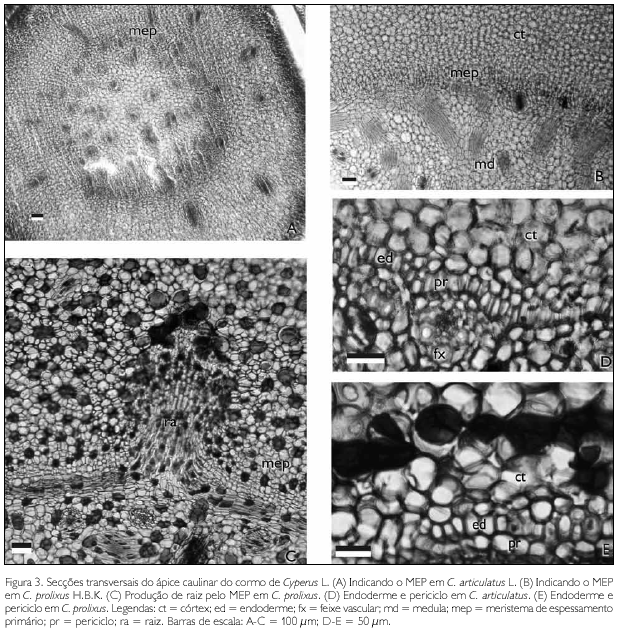
Com o afastamento do ápice caulinar, nas proximidades do primeiro nó, tanto no cormo quanto no rizoma das espécies, é possível observar a diferenciação da endoderme pela presença das estrias de Caspary (Figuras 3D e 3E), originárias da deposição de suberina e de lignina. Subsequente à endoderme, está o periciclo (Figuras 3D e 3E), que mantém a atividade meristemática. Ambos os tecidos substituem o MEP porém, com divisões celulares menos frequentes quando comparados com esse meristema.
No rizoma de C. articulatus, as células corticais, produzidas pelo MEP sofrem lise e originam um aerênquima conspícuo (Figuras 4A e 4B), responsável pela consistência esponjosa do órgão.
]]>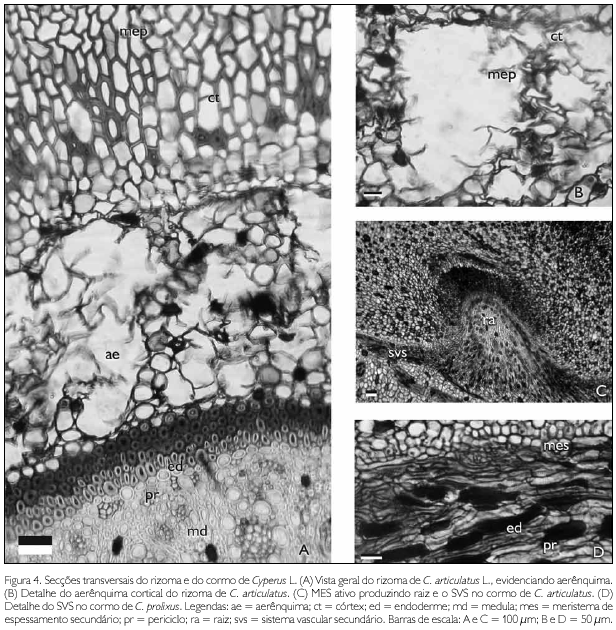
No rizoma de ambas as espécies, o término da diferenciação da endoderme (Figura 4A) coincide com a desaceleração da atividade meristemática do MEP.
Nos cormos das duas espécies, diferentemente dos rizomas, o periciclo não cessa a produção celular com o término da diferenciação da endoderme, assumindo a função de meristema de espessamento secundário (MES) desse órgão (Figura 4C e 4D).
O MES continua produzindo células medulares que permitem o intumescimento do cormo e a produção de novas raízes adventícias. Outra evidência do MES é a produção de células xilemáticas e floemáticas desorganizadas, que não se dispõe em feixes vasculares e representam um sistema vascular secundário na periferia da medula (Figura 4D).
Tanto no cormo quanto no rizoma, nas regiões cortical e medular, foram observados idioblastos com compostos fenólicos (Figuras 4A-4D) e óleo, além de um parênquima amiláceo. No rizoma, tais estruturas secretoras foram menos visualizadas.
Os idioblastos de compostos fenólicos e óleo se destacam pelas dimensões maiores, quando comparados com as células ordinárias (Figura 3C).
DISCUSSÃO
]]> Em Cyperus articulatus e C. prolixus, os caules são dos tipos cormo e rizoma. A presença desses dois tipos de caules simultaneamente não é comum entre as Cyperaceae (Estelita & Rodrigues, 2007), sendo mais frequente a ocorrência de cormos e estolões (Rodrigues & Estelita, 2009), fato este que, nas duas espécies estudadas, evidencia proximidade adaptativa.Os dados morfométricos mostraram que o comprimento do rizoma é relativamente maior que do cormo, assim como o número de nós, indicando que ocorre expansão caulinar contínua nesse órgão. Já o cormo possui maior número de estruturas secretoras e amido, contribuindo para a produção de raízes e demonstrando sua efetiva relação com a reserva de nutrientes e fixação da planta.
Os rizomas de ambas as espécies são perenes, com catafilos, monopodiais e delgados, além de não indicarem abundância de substâncias de reserva, tais como amido, óleos e fenóis, quando comparados ao cormo. Isso ocorre porque o primeiro caule está relacionado exclusivamente à expansão caulinar. Esses rizomas apresentam funções diferentes da maioria das Cyperaceae, os quais, segundo Rodrigues & Estelita (2009), acumulam as funções de expansão caulinar, reserva energética de substâncias orgânicas e de fixação.
Conceição et al. (2008) realizaram levantamento da distribuição de C. articulatus e C. prolixus em herbários no estado do Pará e constataram que o cultivo e a ocorrência eram, geralmente, em locais alagados, entre estes campos e solo alagados, próximo a várzeas, igarapés e lugares paludosos. Essas plantas desenvolvem aerênquimas e rotas internas de aeração, assegurando, assim, alto grau de tolerância à deficiência de oxigênio do solo (Menezes Neto et al., 1995; Vartapetian & Jackson, 1997). O rizoma das espécies estudadas é o órgão de ligação entre diferentes cormos e, consequentemente, espécimes. Dessa forma, a produção de aerênquima observada em C. articulatus demonstra que o rizoma necessita de rotas mais rápidas para a condução de oxigênio, favorecendo processos biológicos ocorrentes em outros órgãos subterrâneos e espécimes.
Sculthorpe (1985) comenta que as plantas aquáticas desenvolvem-se em locais complexos e, por isso, podem apresentar uma alta plasticidade somática. Junk & Piedade (1993) citam que é difícil classificar as espécies da região amazônica nas regiões alagadas, pois elas podem mostrar consideráveis adaptações e viver tanto em terrenos alagados como em terra firme. A ocorrência de tecido aerenquimático no rizoma de C. articulatus evidencia que os indivíduos coletados ainda apresentam características de plantas de ambiente alagado. As amostras de C. prolixus apresentam adaptação à terra firme, pois o tecido parenquimático, altamente influenciado pelas condições ambientais nas quais a planta vive, pode ou não se diferenciar em aerênquima.
Em C. articulatus e C. prolixus, o cormo é perene, com catafilo, intumescido e possui a função de promover a propagação vegetativa, devido à presença de botões apicais, de abundantes substâncias de reserva energética, na forma de amido, óleos e fenóis, fazendo esse o órgão com maior quantidade de compostos orgânicos, e do MES, que provê suporte estrutural para o crescimento e resistência às mudanças ambientais. Tradicionalmente, essas funções, em Cyperaceae, eram atribuídas ao rizoma (Metcalfe, 1971), sendo que, recentemente, foram considerados outros tipos de caule, tal como o cormo, com essas funcionalidades (Estelita & Rodrigues, 2007).
A quantidade de substância de reserva no cormo está relacionada diretamente com a produção de respiração anaeróbica durante períodos prolongados de alagamentos (Taiz & Zeiger, 2004), contribuindo para a manutenção da latência ou sobrevivência em períodos de anoxia crônica.
A presença de MEP e MES em espécies de Cyperus ocorrentes no Brasil foi previamente demonstrada em trabalhos como os de Rodrigues & Estelita (2002). Geralmente, tais tecidos estão relacionados com a produção de tecidos condutores, parênquima e raízes adventícias, concomitante ao crescimento apical (Rudall, 1991).
Em C. articulatus e C. prolixus, o MEP encontra-se ativo tanto no rizoma quanto no cormo, com função distinta entre os dois órgãos, já que no cormo o MEP pode produzir raízes adventícias e no rizoma, não.
Uma explicação à atividade observada no MEP está relacionada com as funções do cormo e do rizoma. O primeiro é responsável pela propagação, necessitando de substâncias de reserva energética, como amido, e fixação ao substrato, enquanto o segundo se relaciona com a expansão, não havendo necessidade de abundância de reserva e de fixação.
]]> Nas espécies estudadas, ficou evidente que, no cormo e no rizoma, o MEP é o responsável direto pela formação da endoderme e do periciclo, estando esses três tecidos intimamente conectados em cronologia, ou seja, o meristema e os tecidos maduros oriundos do desenvolvimento do corpo vegetal. Essa análise diverge das expostas por Menezes et al. (2005), que relataram a inexistência do tecido MEP em monocotiledôneas, mas sim a precoce ocorrência da endoderme e do periciclo desde o ápice caulinar, gerando um espessamento primário.O MES no cormo de C. articulatus e C. prolixus é respaldado pela presença do sistema vascular secundário. Esse sistema, segundo Rodrigues & Estelita (2002), é constituído por xilema e floema não organizados em feixes.
Rudall (1991) e Rodrigues & Estelita (2002) relataram a possibilidade de que a ocorrência simultânea do MEP e do MES, na mesma espécie, seja uma plesiomorfia em relação à ocorrência apenas do MEP entre as monocotiledôneas. Por conseguinte, as espécies estudadas são de transição nesse caractere evolutivo, visto que, na mesma espécie, pode ocorrer um caule com MEP e MES, o cormo, e outro só com o MEP o rizoma. Logo, supõe-se que os rizomas perderam o MES, enquanto os cormos os mantiveram.
Tradicionalmente, as raízes adventícias de caules aéreos são originárias das regiões de nós do caule e tendem a crescer na direção oposta à superfície, visando à fixação e absorção (Bell, 2008). Em C. articulatus e C. prolixus, as raízes são exclusivamente adventícias do cormo subterrâneo, a partir de nós e entrenós e crescendo em todas as direções.
O modelo observado de formação de raízes adventícias nas espécies estudadas tem relação direta com o MES, uma vez que são produtos da ação desse meristema. Como o MES ocorre apenas nos cormos, é a partir dele que se desenvolvem estas raízes, diferentemente do rizoma, no qual não há MES. Em adição, o MES envolve todo o interior do cormo, produzindo, desse modo, raízes em todas as direções durante sua atuação.
CONCLUSÃO
Os resultados desta pesquisa permitem considerar os caules de Cyperus articulatus e de C. prolixus como complexos, sendo constituídos por rizomas e cormos. Entre estes, o rizoma mostra-se mais adaptado à expansão do corpo vegetativo, enquanto o cormo, à reserva energética, propagação, fixação e absorção.
Nas duas espécies estudadas, os MEP e MES são fundamentais na formação do caule. O MEP ocorre no rizoma e no cormo, não produzindo raízes no primeiro, já o MES encontra-se apenas no cormo, produzindo aumento considerável do diâmetro deste caule em relação ao rizoma.
]]> AGRADECIMENTOS
Os autores agradecem ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), pela concessão da bolsa no Programa de Pós-graduação em Botânica ao primeiro autor; ao Programa de Pesquisa em Biodiversidade (PPBio), pelo apoio a este projeto; e ao engenheiro florestal João B. da Silveira, pelas ilustrações botânicas.
REFERÊNCIAS
ALVES, M. V., A. C. ARAÚJO, A. N. PRATA, F. VITTA, S. HEFLER, R. TREVISAN, A. S. B. GIL, S. MARTINS & W. THOMAS, 2009. Diversity of Cyperaceae in Brazil. Rodriguésia 60(4): 771-782.
BELL, A. D., 2008. Plant form: An illustrated guide to flowering plant morphology. Timber Press, Portland, London.
BRAGA, M., 1977. Anatomia foliar de Bromeliaceae da Campina. Acta Amazonica 7(suplemento): 1-74.
CONCEIÇÃO, C. C. C., A. B. SILVA, M. G. C. MOTA & M. S. F. REDIG, 2008. Distribuição geográfica e coleta de germoplasma de priprioca (Cyperus spp.) no estado do Pará. In: R. C. V. POTIGUARA & M. G. B. ZOGHBI (Orgs.): Priprioca: um recurso aromático do Pará: 113-129. Museu Paraense Emílio Goeldi/Universidade do Estado do Pará (Coleção Adolpho Ducke), Belém.
ESTELITA, M. E. M. & A. C. RODRIGUES, 2007. Subsídios estruturais à caracterização do sistema caulinar em Cyperaceae. Revista Brasileira de Botânica 30(3): 401-409.
GONÇALVES, E. G. & H. LORENZI, 2007. Morfologia vegetal: organografia e dicionário ilustrado de morfologia das plantas vasculares: 21-24. Instituto Plantarum de Estudos da Flora, Nova Odessa.
JOHANSEN, D. A., 1940. Plant microtechnique. McGraw Hill, New York.
JUDD, S. W., C. S. CAMPBELL, E. A. KELLOG, P. F. STEVENS & M. J. DONOGHUE, 2009. Sistemática vegetal: um enfoque filogenético: 3. ed.: 1-632. Artmed, Porto Alegre.
JUNK, W. J. & M. T. F. PIEDADE, 1993. Herbaceous plants of the Amazon floodplain near Manaus: Species diversity and adaptations to the flood pulse. Amazoniana 12(3-4): 467-484.
MENEZES, N. L., D. C. SILVA, R. C. O. ARRUDA, G. F. MELO-DE-PINNA, V. A. CARDOSO, N. M. CASTRO, V. L. SCATENA & E. SCREMIN-DIAS, 2005. Meristematic activity of the endodermis and the pericycle in the primary thickening in monocotyledons. Considerations on the "PTM". Anais da Academia Brasileira de Ciências 77(2): 259-274.
MENEZES NETO, M. A., J. D. ALVES & L. E. M. OLIVEIRA, 1995. Anaerobic metabolism of Euterpe oleracea. II-Plant tolerance mechanism to anoxia. Revista Brasileira de Fisiologia Vegetal 7(1): 47-51.
METCALFE, C. R., 1971. Anatomy of the monocotyledons: Cyperaceae. v. 5: 15-44. Clarendon Press, Oxford.
POTIGUARA, R. C. V. & P. P. SANTOS, 2008. Morfo-anatomia dos órgãos aéreos e subterrâneos de Cyperus articulatus L. (Cyperaceae Juss.). In: R. C. V. POTIGUARA & M. G. B. ZOGHBI (Orgs.): Priprioca: um recurso aromático do Pará: 25-41. Museu Paraense Emílio Goeldi/Universidade do Estado do Pará (Coleção Adolpho Ducke), Belém.
RODRIGUES, A. C. & M. E. M. ESTELITA, 2002. Primary and secondary development of Cyperus giganteus Vahl rhizome (Cyperaceae). Revista Brasileira de Botânica 25(3): 251-258.
RODRIGUES, A. C. & M. E. M. ESTELITA, 2009. Morphoanatomy ofthe stem in Cyperaceae. Acta Botanica Brasilica 23(3): 889-901.
RUDALL, P., 1991. Lateral meristems and stem thickening growth in monocotyledons. The Botanical Review 57(2): 150-163.
SCULTHORPE, C. D., 1985. The biology of aquatic vascular plants. Koeltz Scientific Books, Kónigstein.
TAIZ, L. & E. ZEIGER, 2004. Fisiologia vegetal. 3. ed. Artmed, Porto Alegre.
VARTAPETIAN, B. & M. B. JACKSON, 1997. Plant adaptations to anaerobic stress. Annals of Botany 79(suplemento 1): 3-20.
ZOGHBI, M. G. B., E. H. A. ANDRADE, L. M. M. CARREIRA & A. E. S. ROCHA, 2008. Comparison of the main components of the essential oils of priprioca: Cyperus articulatus var. articulatus L., C. articulatus var. nodosus L., C. prolixus Kunth and C. rotundus L. The Journal of Essential Oil Research 20(1): 42-46.
 Autor para correspondência:
Autor para correspondência:
Pedro Paulo dos Santos.
Museu Paraense Emílio Goeldi/Ministério da Ciência,
Tecnologia e Inovação. ]]>
Coordenação de Botânica,
Laboratório de Anatomia Vegetal.
Av. Perimetral, 1901 – Terra Firme. Belém, PA, Brasil.
CEP 66077-530.
(santos_pp@yahoo.com.br)
Recebido em 11/02/2011
Aprovado em 01/04/2012
Responsabilidade editorial: Anna Luiza IIkiu-Borges
]]>